Identification of amino-terminal region of adiponectin
as a physiologically functional domain
Adiponectin is an abundant adipose-specific
protein, which acts as an anti-diabetic, anti-atherogenic,
and anti-inflammatory adipokine. Although recent advances
in the field of adiponectin have been made by the identification
of adiponectin receptors and by the understanding about
relationship between its multimerization and functions,
detailed molecular background remains unclear. Our established
anti-human adiponectin antibodies, ANOC 9103 and ANOC 9104,
blocked some adiponectin functions such as the growth inhibition
of B-lymphocytes on stromal cells and the inhibition of
acetylated LDL uptake in macrophages, suggesting that they
may recognize important functional regions of adiponectin.
As a result of epitope mapping based on the ability to bind
to the deleted adiponectin mutants, we identified that these
antibodies recognize amino-terminal region of adiponectin
before the beginning of the collagen-like domain. Notably,
a peptide fragment (DQETTTQGPGVLLPLPKGACTGWMA) corresponding
to amino acid residues 17-41 of human adiponectin could
bind to restricted types of cells and block adiponectin-induced
cyclooxygenase-2 gene expression and prostaglandin E(2)
production in MS-5 stromal cells. Moreover, the deletion
of its amino-terminal region reduced the abilities to inhibit
not only collagen-induced platelet aggregation but also
diet-induced hepatic steatosis. These data indicate that
amino-terminal region of adiponectin is a physiologically
functional domain and that a novel receptor, which recognizes
amino-terminal region of adiponectin, may exist on some
types of cells. Further investigations will contribute to
the understanding of molecular mechanisms about adiponectin
functions as well as to the designing of novel strategies
for the treatment of patients with insulin-resistance, vascular
dysfunction, and chronic inflammation.
Ujiie H,
et al. J Cell Biochem. 2006 Jan 11; [Epub ahead of
print]
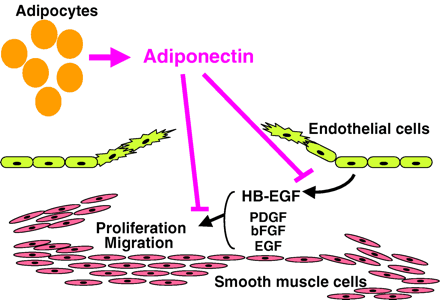
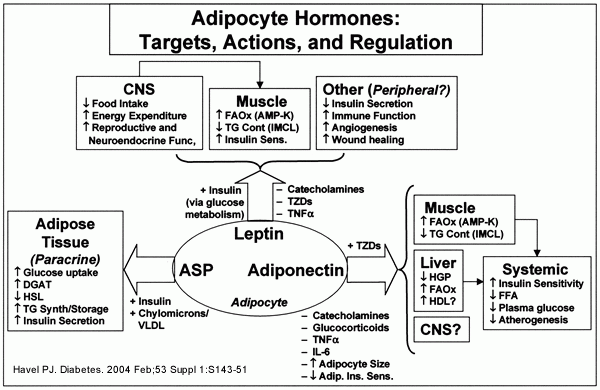
Model illustrating how adiponectin exerts a preventive effect
on vascular stenosis in the injured artery. J.
Biol. Chem., Vol. 277, Issue 40, 37487-37491, October 4, 2002
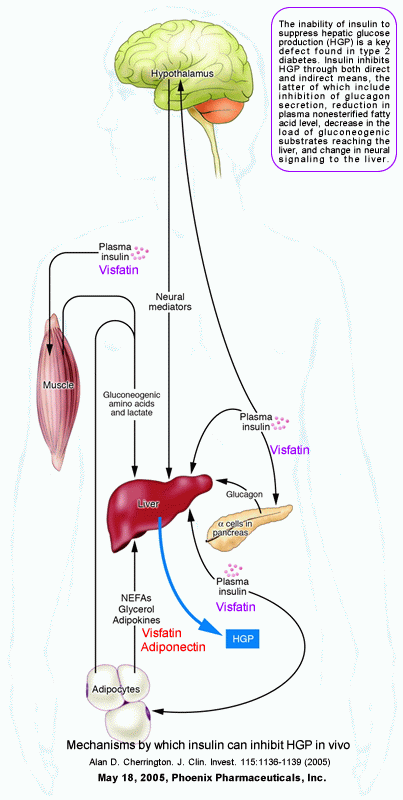
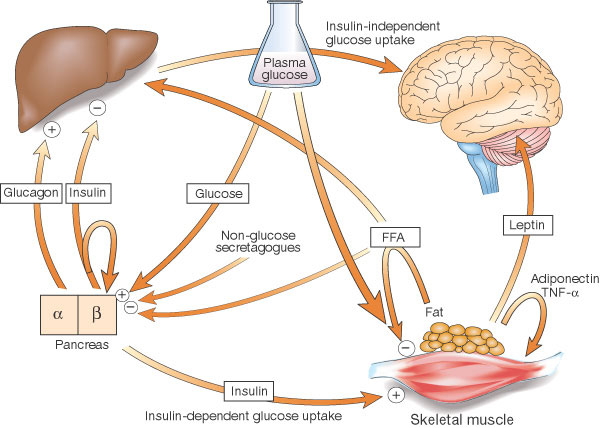
Mechanisms by which insulin can inhibit HGP in vivo
A hypothetical model for the secretion and action of adiponectin.
The synthesis and secretion of adiponectin is increased
by activation of the nuclear receptor PPAR-gamma, and reduced
by caloric excess, presumably associated with leptin deficiency
or resistance. Once released, adiponectin can directly increase
fatty-acid transport, oxidation and dissipation in skeletal
muscle, reducing the levels of intramyocellular lipids,
thus improving insulin signaling. The protein can also increase
the sensitivity of the hepatocyte to insulin, either through
a direct action, or indirectly by lowering circulating lipids
due to its action on muscle. Thus, administration of adiponectin
can result in improved insulin sensitivity and glucose tolerance,
and can correct hyperglycemia associated with obesity.
Saltiel AR. You are what you secrete.
Nat Med. 2001 Aug;7(8):887-8
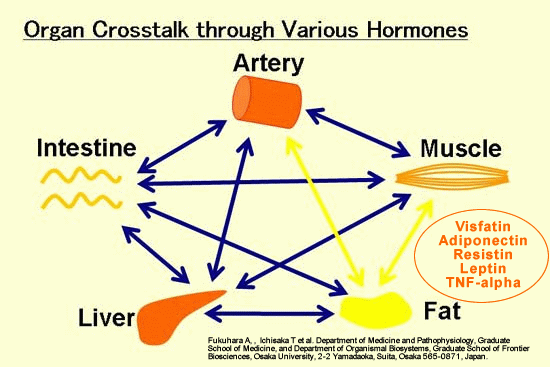
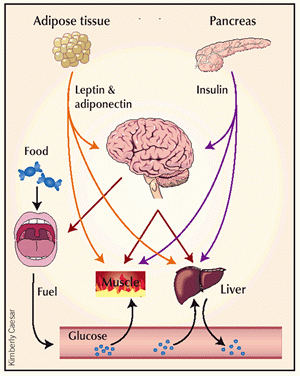
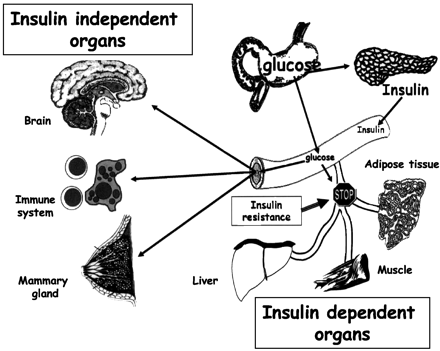
Cross-talk between tissues
in the regulation of glucose metabolism. Insulin is secreted
from the ß-cells of the pancreas in response to elevations
in plasma glucose. The hormone decreases glucose production
from the liver, and increases glucose uptake, utilization
and storage in fat and muscle. The fat cell is important
in metabolic regulation, releasing FFAs that reduce glucose
uptake in muscle, insulin secretion from the ß-cell, and
increase glucose production from the liver. The fat cell
can also secrete 'adipokines' such as leptin, adiponectin
and TNF, which regulate food intake, energy expenditure
and insulin sensitivity.
ALAN R. SALTIEL* AND C. RONALD KAHN.
Nature414, 799 - 806 (2001); doi:10.1038/414799a
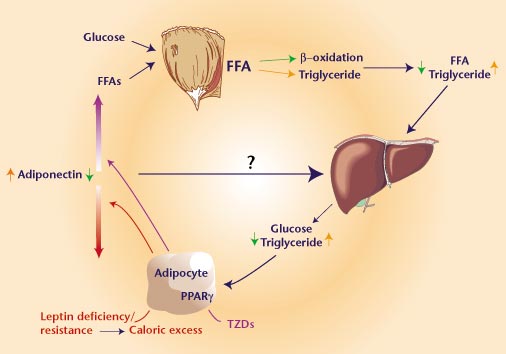
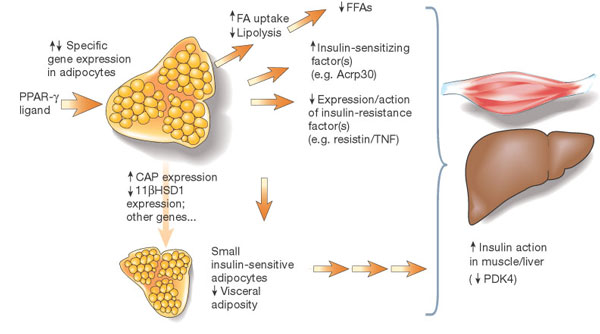
Potential mechanisms of
insulin sensitization by PPAR-gamma ligands. The receptor
PPAR-gamma is predominantly expressed in adipose tissue.
Ligand interactions with the receptor mediate specific changes
in adipose gene expression. Altered expression of adipose
genes such as fatty-acid transporter 1 may contribute to
reduced production of free fatty acids (FFAs), which, in
turn, is predicted to have insulin-sensitizing effects in
muscle and liver. Changes in expression of other genes such
as CAP or 11ßHSD1 may contribute to locally increased insulin
action in adipose tissue and/or reduced visceral adiposity.
Altered expression of circulating factors including TNF-,
resistin and Acrp30 is also likely to indirectly mediate
increased action of insulin in liver or muscle and glucose
utilization; suppression of PDK4 activity in muscle is an
example of one (probably indirect) effect. DAVID E. MOLLER.
Nature
414, 821 - 827 (2001); doi:10.1038/414821a
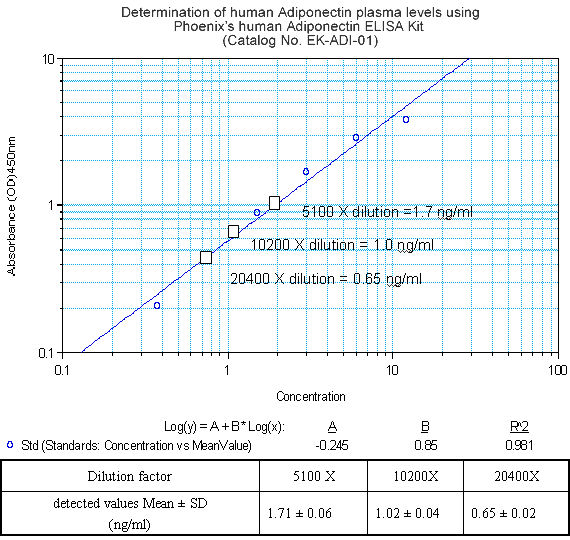
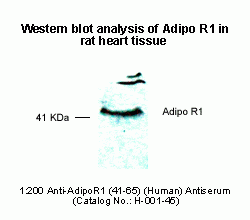
Western Blot Analysis of Adiponectin
Adiponectin Receptor 1 Western
Blot Analysis
Antibody for Adiponectin Receptor 1
Immunohistochemistry
The four modified lysines (residues 68, 71, 80, and
104) in the collagenous domain of adiponectin are conserved
across all of the species investigated. The mouse, human,
bovine, monkey, and dog adiponectin sequences relate to GenBankTM/EBI
accession numbers BAB22597, NP_004788, AAK58902, AAK92202,
and AAL09702, respectively. The four modified lysines and
their surrounding motifs are shaded. Wang
Y, et al. J Biol Chem. 2002 May 31;277(22):19521-9.
Epub 2002 Mar 23
Adiponectin acts in the brain to decrease body weight
Adiponectin (ADP) is an adipocyte hormone
involved in glucose and lipid metabolism. We detected a
rise in ADP in cerebrospinal fluid after intravenous (i.v.)
injection, consistent with brain transport. In contrast
to leptin, intracerebroventricular (i.c.v.) administration
of ADP decreased body weight mainly by stimulating energy
expenditure. Full-length ADP, mutant ADP with Cys39 replaced
with serine, and globular ADP were effective, whereas the
collagenous tail fragment was not. Lepob/ob mice were especially
sensitive to i.c.v. and systemic ADP, which resulted in
increased thermogenesis, weight loss and reduction in serum
glucose and lipid levels. ADP also potentiated the effect
of leptin on thermogenesis and lipid levels. While both
hormones increased expression of hypothalamic corticotropin-releasing
hormone (CRH), ADP had no substantial effect on other neuropeptide
targets of leptin. In addition, ADP induced distinct Fos
immunoreactivity. Agouti (Ay/a) mice did not respond to
ADP or leptin, indicating the melanocortin pathway may be
a common target. These results show that ADP has unique
central effects on energy homeostasis.
Nature Medicine, Published online:
11 April 2004, doi:10.1038/nm1029
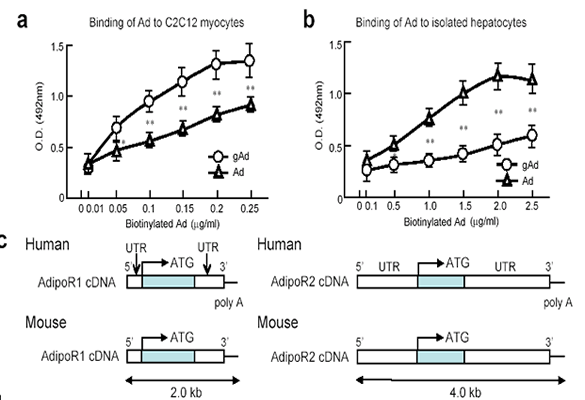
Cloning of adiponectin receptors that mediate
antidiabetic metabolic effects
Adiponectin (also known
as 30-kDa adipocyte complement-related protein; Acrp30)
is a hormone secreted by adipocytes that acts as an antidiabetic
and anti-atherogenic adipokine. Levels of adiponectin in
the blood are decreased under conditions of obesity, insulin
resistance and type 2 diabetes. Administration of adiponectin
causes glucose-lowering effects and ameliorates insulin
resistance in mice. Conversely, adiponectin-deficient mice
exhibit insulin resistance and diabetes. This insulin-sensitizing
effect of adiponectin seems to be mediated by an increase
in fatty-acid oxidation through activation of AMP kinase
and PPAR-alpha. Here we report the cloning of complementary
DNAs encoding adiponectin receptors 1 and 2 (AdipoR1 and
AdipoR2) by expression cloning. AdipoR1 is abundantly expressed
in skeletal muscle, whereas AdipoR2 is predominantly expressed
in the liver. These two adiponectin receptors are predicted
to contain seven transmembrane domains, but to be structurally
and functionally distinct from G-protein-coupled receptors.
Expression of AdipoR1/R2 or suppression of AdipoR1/R2 expression
by small-interfering RNA supports our conclusion that they
serve as receptors for globular and full-length adiponectin,
and that they mediate increased AMP kinase and PPAR-alpha
ligand activities, as well as fatty-acid oxidation and glucose
uptake by adiponectin.
Yamauchi T, et al. Nature.
2003 Jun 12;423(6941):762-9
Plasma Adiponectin Increases Postprandially
in Obese, but not in Lean, Subjects
OBJECTIVE: We investigated
the acute responses of plasma adiponectin levels to a test
meal in lean and obese subjects. RESEARCH METHODS AND PROCEDURES:
We studied 13 lean and 11 obese subjects after a 10-hour
overnight fast. Glucose, insulin, and adiponectin concentrations
were measured at baseline and 15, 30, 60, 120, and 180 minutes
after a fixed breakfast. RESULTS: At baseline, fasting adiponectin
concentrations were lower in the obese group vs. the lean
group [mean (95% confidence interval): 2.9 (2.1 to 4.1)
micro g/mL vs. 8.6 (6.5 to 11.3) micro g/mL], but rose 4-fold
postprandially in the obese group, reaching a peak at 60
minutes [baseline: 2.9 (2.1 to 4.1) micro g/mL vs. 60 minutes:
12.1 (8.5 to 17.4) micro g/mL; p< 0.0001] and remaining
elevated for the remainder of the study. There were no postprandial
changes in plasma adiponectin concentrations in lean subjects.
DISCUSSION: This increase of adiponectin concentrations
in obese individuals might have important beneficial effects
on postprandial glucose and lipid metabolism and might be
viewed as a mechanism for maintaining normal glucose tolerance
in those who are obese and insulin resistant.
English PJ, et al. Obes
Res. 2003 Jul;11(7):839-44
Ghrelin and adipose tissue regulatory peptides:
effect of gastric bypass surgery in obese humans
Presently surgery is the
most effective way to obtain a controlled weight reduction
in morbidly obese patients. Roux-en-Y gastric bypass (RYGBP)
surgery is effective and used worldwide, but the exact mechanism
of action is unknown. The effect of RYGBP on ghrelin, insulin,
adiponectin, and leptin levels was investigated in 66 obese
subjects; mean weight 127 kg (range, 96-195 kg) and mean
body mass index (BMI) 45 kg/m(2) (range, 33-64) before and
after surgery. Ghrelin levels were also compared in 10 nonoperated
and 10 operated obese, BMI-matched women. RYGBP resulted
in 22% and 30% weight loss at 6 and 12 months, respectively.
Ghrelin increased by 44% and 62% and adiponectin by 36%
and 98%, but insulin declined by 57% and 62% and leptin
by 60% and 64%. The changes were all related to the reduction
in BMI. In addition, ghrelin and insulin were inversely
correlated at all time points as were changes of the peptides
at 12 months (F = 4.9, P = 0.031), independent of the change
in BMI. No evidence for RYGBP surgery per se having an effect
on ghrelin levels, independent of weight loss, was obtained.
The profound changes in the regulatory peptides are likely
to reflect the new state of energy balance achieved. A close
inverse association between ghrelin and insulin was observed,
supporting an important role for ghrelin in glucose homeostasis.
Holdstock C, et al. J
Clin Endocrinol Metab. 2003 Jul;88(7):3177-83
The fat-derived hormone adiponectin alleviates
alcoholic and nonalcoholic fatty liver diseases in mice
Adiponectin has recently
been shown to be a promising candidate for the treatment
of obesity-associated metabolic syndromes. Replenishment
of recombinant adiponectin in mice can decrease hyperglycemia,
reverse insulin resistance, and cause sustained weight loss
without affecting food intake. Here we report its potential
roles in alcoholic and nonalcoholic fatty liver diseases
in mice. Circulating concentrations of adiponectin decreased
significantly following chronic consumption of high-fat
ethanol-containing food. Delivery of recombinant adiponectin
into these mice dramatically alleviated hepatomegaly and
steatosis (fatty liver) and also significantly attenuated
inflammation and the elevated levels of serum alanine aminotransferase.
These therapeutic effects resulted partly from the ability
of adiponectin to increase carnitine palmitoyltransferase
I activity and enhance hepatic fatty acid oxidation, while
it decreased the activities of two key enzymes involved
in fatty acid synthesis, including acetyl-CoA carboxylase
and fatty acid synthase. Furthermore, adiponectin treatment
could suppress the hepatic production of TNF-alpha and plasma
concentrations of this proinflammatory cytokine. Adiponectin
was also effective in ameliorating hepatomegaly, steatosis,
and alanine aminotransferase abnormality associated with
nonalcoholic obese, ob/ob mice. These results demonstrate
a novel mechanism of adiponectin action and suggest a potential
clinical application of adiponectin and its agonists in
the treatment of liver diseases.
Xu A, et al. J Clin Invest. 2003 Jul;112(1):91-100
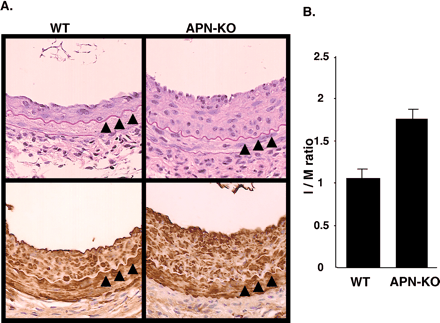
Intimal thickening of injured arteries
in adiponectin-deficient mice. A, representative hematoxylin
and eosin-stained sections (upper panel) and -smooth muscle-actin
immunostained sections (lower panel) of femoral arteries of
WT and adiponectin knockout (APN-KO) male mice (8-10 weeks
old) harvested at 3 weeks after injury. Arrows indicate internal
elastic lamina. B, I/M ratio in the injured arteries of WT
(n = 7, ±S.E.) and APN-KO (n = 5, ±S.E.) mice (p < 0.01,
t test). J. Biol. Chem., Vol. 277, Issue
40, 37487-37491, October 4, 2002
Figure 2
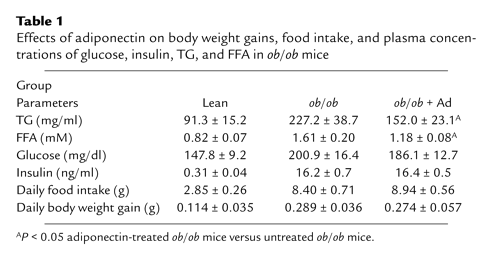
Effects of adiponectin treatment on food intake, body weight
gains, and circulating levels of glucose and insulin. (a)
Serum adiponectin levels in mice fed with LC diet, LE diet,
or LE diet plus adiponectin infusion (LE + Ad), as described
in the text (n = 5). Serum samples were collected at
different times following adiponectin treatment. (b)
Food intake in LE diet mice treated with or without adiponectin
(n = 6). (c) Average daily body weight gains
over 2 weeks of adiponectin treatment (n = 6). (d)
Fasting glucose levels over adiponectin treatment period (n
= 6). (e) Fasting plasma insulin levels over adiponectin
treatment period (n = 6). Note that adiponectin and
alcohol treatment also have no obvious effects on plasma levels
of insulin and glucose in the fed state (data not shown).
*P < 0.001 for adiponectin-treated group versus
other two groups.
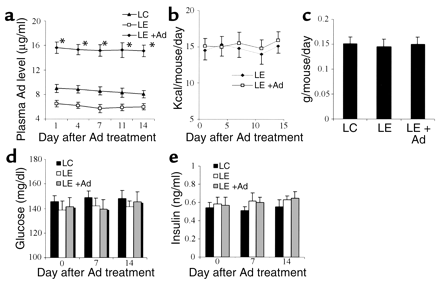
Figure 3
Adiponectin treatment ameliorates alcohol-induced dyslipidemia
and hepatic abnormality in mice. Serum samples were collected
after 5 weeks of LC diet, LE diet, or LE + Ad diet in the
last 2 weeks. Serum levels of TG (a) and FFA (b),
liver-to-body weight ratio (c), hepatic TG contents
(d), and plasma ALT levels (e) were determined
at necropsy (n = 5). #P < 0.05,
##P < 0.01 for LC-treated mice versus
LE-treated mice; *P < 0.05, **P < 0.01
for LE + Ad–treated mice versus LE-treated mice.
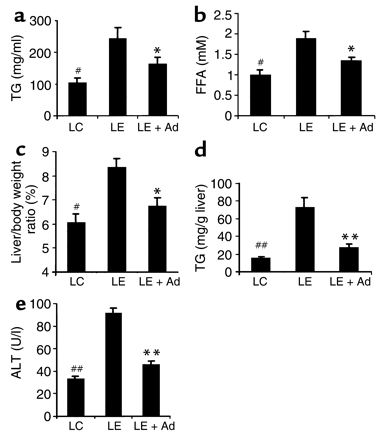
Figure 5
Adiponectin treatment decreases alcohol-induced elevation
of hepatic TNF-
expression and its plasma concentrations. (a) Total
RNA from livers of mice treated with LC diet, LE diet, or
LE + Ad diet was extracted and subjected to Northern blot
analysis. (b) The results from a were quantified
by PhosphorImaging (n = 5). All RNA levels are expressed
relative to untreated LC pair-fed controls, after being normalized
against the abundance of 18S RNA. (c) Plasma concentrations
of TNF-
as measured at necropsy (n = 5). **P < 0.01
for LE + Ad–treated mice versus mice receiving LE diet
alone.
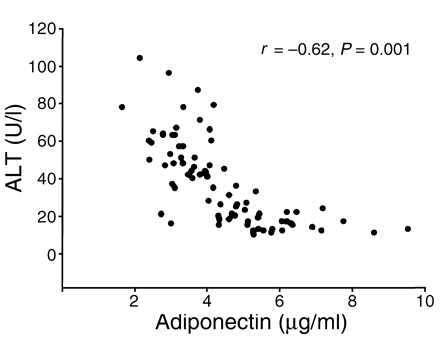
Relationship between plasma
levels of adiponectin and ALT in 90 morbidly obese Chinese
individuals (45 male and 45 female). r, Spearman correlation
Hydroxylation and glycosylation
of the four conserved lysine residues in the collagenous domain
of adiponectin. Potential role in the modulation of its insulin-sensitizing
activity. Wang Y, et al. J Biol Chem. 2002 May 31;277(22):19521-9.
Epub 2002 Mar 23
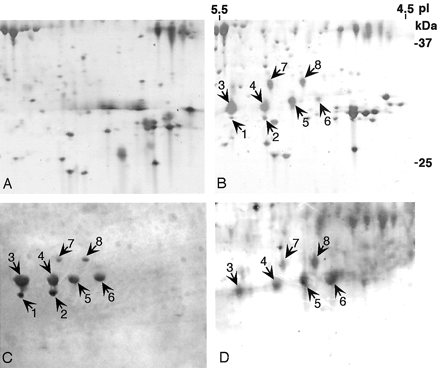
Endogenous adiponectin
secreted by 3T3-L1 adipocytes exists as eight isoforms.
The medium from subconfluent 3T3-L1 preadipocytes (A)
or adipocytes at day 8 after induction of differentiation
(B) was collected and concentrated as described under
"Experimental Procedures." 50 µg of proteins from each
sample was separated by two-dimensional gel electrophoresis
and visualized by silver staining. The proteins preferentially
secreted by adipocytes are denoted by numbered arrows.
Secretory proteins from adipocytes separated by two-dimensional
gel electrophoresis as described above were detected using
rabbit anti-adiponectin antibody (C) or the Immu-Blot
carbohydrate detection kit (D). Note that all eight
indicated proteins are immunoreactive with anti-adiponectin
antibody. Six of the eight isoforms (spots 3-8) of
adiponectin are glycosylated.
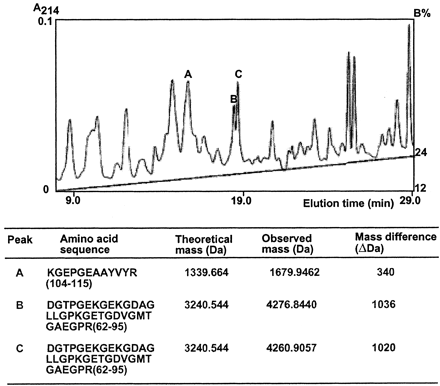
Fractionation and characterization of the tryptic peptides
of adiponectin by RP-HPLC, MALDI-TOF-MS, and amino acid sequencing.
All of the glycosylated isoforms of adiponectin separated
by two-dimensional gel electrophoresis were excised from the
gels, pooled, and digested with trypsin. The tryptic peptide
mixture was separated by RP-HPLC. Each fraction was collected
and analyzed by MALDI-TOF-MS. The three fractions containing
peptides with masses of 1679, 4276, and 4260 Da
are denoted as A-C, respectively. The table
shows the amino acid sequences, the experimentally observed
masses, the theoretical masses, and the mass differences for
these three peptides.
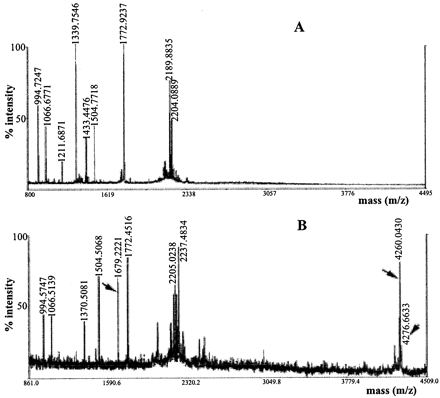
MALDI-TOF mass spectra
of tryptic peptide mixtures derived from different isoforms
of adiponectin. Isoforms 1 (A) and 3 (B)
of adiponectin secreted by adipocytes were in-gel digested
with trypsin, and the tryptic mixtures were analyzed by MALDI-TOF-MS.
Note that the three peptides (with masses of 1679, 4260, and
4276 Da) denoted by arrows were reproducibly observed
in all of the glycosylated isoforms (isoforms 3-8) produced
by both adipocytes and COS-7 cells, and not in the two unglycosylated
isoforms (isoforms 1 and 2) or the bacterially produced
adiponectin.
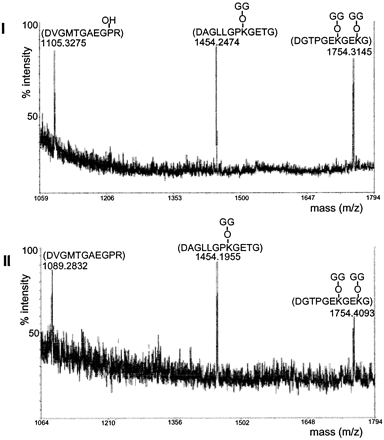
MALDI-TOF mass spectra
of peptide mixtures from Asp-N-digested peptides B and C. Peptides
B (panel I) and C (panel II) separated in Fig.
3 were further digested with Asp-N and then analyzed by MALDI-TOF-MS.
The peptide sequences and the potential modifications assigned
to each mass are indicated above each peak. Note that the
assignment of Pro94 as hydroxylated proline was
also confirmed by amino acid analysis. GG, glucosylgalactosyl.
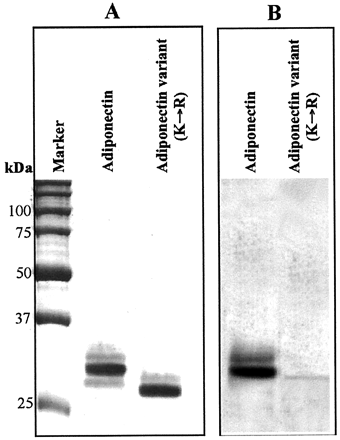
Expression and carbohydrate
detection of the FLAG-tagged Lys-to-Arg adiponectin variant.
COS-7 cells were transfected with pcDNA-Ad-F or pcDNA-Ad(KR)-F.
48 h later, FLAG-tagged adiponectin or the Lys-to-Arg
adiponectin variant was purified from the cell culture medium.
1 µg of protein from each sample was separated by 15%
SDS-PAGE and stained with Coomassie Brilliant Blue R-250 (A)
or detected with the Immu-Blot glycoprotein detection kit
(B). Note that the majority of glycosylation was abolished
in the Lys-to-Arg adiponectin variant.
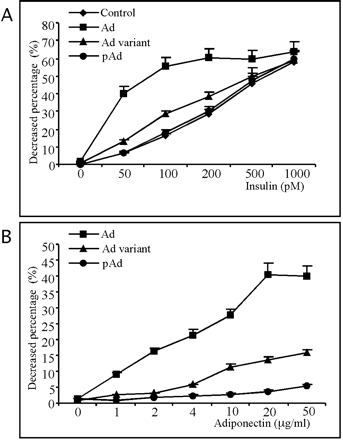
Adiponectin can enhance the action of insulin to inhibit
hepatic glucose production
Effect of adiponectin and adiponectin variants on insulin-induced
inhibition of glucose production in primary rat hepatocytes.Upper panel, inhibition of hepatic glucose production
following treatment with increasing amounts of insulin in
the absence or presence of 20 µg/ml adiponectin (Ad)
or Lys-to-Arg adiponectin variant (Ad variant) generated
from COS-7 cells or 20 µg/ml bacterially produced adiponectin
(pAd); lower panel, inhibition of hepatic glucose
production following treatment with 50 pM
insulin plus increasing amounts of adiponectin or the Lys-to-Arg
adiponectin variant generated from COS-7 cells or bacterially
produced adiponectin. The results are represented as decreased
percentage of glucose production relative to the untreated
cells and as means ?nbsp;S.D. (n = 4).
Adipose tissue can have
a substantial impact on systemic glucose homeostasis through
production of bioactive molecules. We examined serum concentrations
of adipocyte-derived secreted proteins with postulated roles
in obesity and insulin action. ACRP30 (30-kDa adipocyte complement-related
protein)/adiponectin concentrations in the obese Jnk1-/-
mice were significantly higher than in Jnk1+/+
controls (Fig.
2e). In contrast, the concentrations of resistin were
lower in Jnk1-/- mice than in Jnk1+/+
animals (Fig.
2f). Because recent studies have indicated a role for
adiponectin as a mediator of fatty-acid oxidation and hepatic
insulin sensitivity and resistin is postulated to have a role
in insulin resistance, these alterations could affect systemic
insulin sensitivity.
HIROSUMI J. et al. Nature
420, 333 - 336 (21 November 2002); doi:10.1038/nature01137
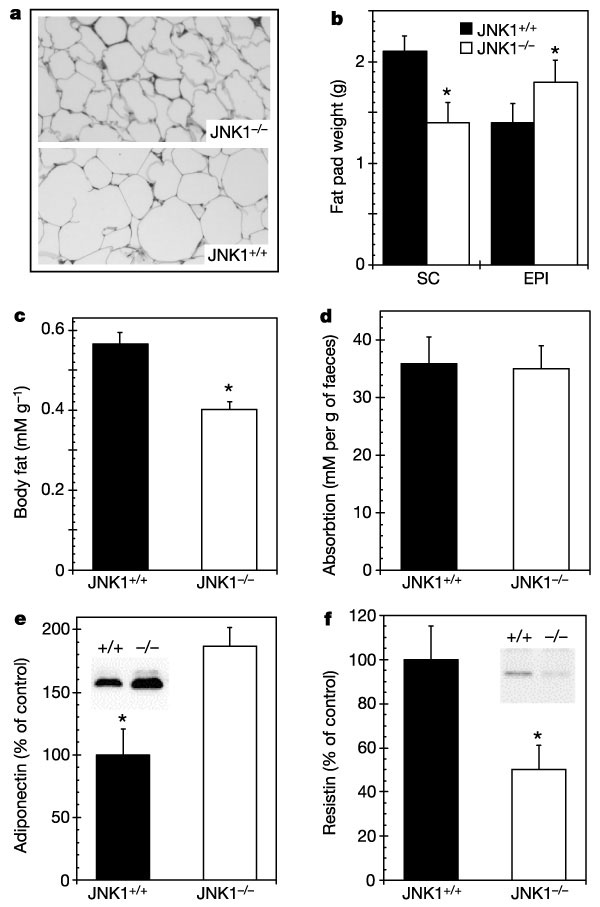
Fig. 2 Adipose tissue morphology
and adiposity in Jnk1-/- mice and wild-type
controls. a, b, Histological sections of epididymal
fat pads (original magnification 50)
(a) and subcutaneous (SC) and epididymal (EPI) fat
pad weights (b) of 16-week-old male Jnk1-/-
and Jnk1+/+ mice (n = 3 in a,
n = 9 in b). c?b>f, Total body composition
(c), faecal lipid content (d), serum adiponectin
concentration (e) and resistin concentration (f)
in Jnk1-/- and Jnk1+/+
mice. Representative immunoblots are shown in insets. Total
carcass lipid analysis was performed25 to determine
fat mass of individual mice (n = 6 in each group).
Asterisk, statistical significance (P < 0.05) in
a two-tailed Student t-test comparing Jnk1+/+
and Jnk1-/- mice.