Angiopoietin-related
Growth Factor (AGF) /ANGIOPOIETIN-LIKE 6 (ANGPTL6)
A
new hepatocyte-derived circulating factor that counteracts obesity
and related insulin resistance

Angiopoietin-related growth factor (AGF)
promotes epidermal proliferation, remodeling, and regeneration
We report here the identification of an angiopoietin-related
growth factor (AGF). To examine the biological function of AGF
in vivo, we created transgenic mice expressing AGF in epidermal
keratinocytes (K14-AGF). K14-AGF mice exhibited swollen and
reddish ears, nose and eyelids. Histological analyses of K14-AGF
mice revealed significantly thickened epidermis and a marked
increase in proliferating epidermal cells as well as vascular
cells in the skin compared with nontransgenic controls. In addition,
we found rapid wound closure in the healing process and an unusual
closure of holes punched in the ears of K14-AGF mice. Furthermore,
we observed that AGF is expressed in platelets and mast cells,
and detected at wounded skin, whereas there was no expression
of AGF detected in normal skin tissues, suggesting that AGF
derived from these infiltrated cells affects epidermal proliferation
and thereby plays a role in the wound healing process. These
findings demonstrate that biological functions of AGF in epidermal
keratinocytes could lead to novel therapeutic strategies for
wound care and epidermal regenerative medicine.
Oike Y, et al. Proc Natl Acad
Sci U S A. 2003 Aug 5;100(16):9494-9
Angiopoietin-related growth factor (AGF) promotes angiogenesis
We report here the identification of angiopoietin-related growth
factor (AGF) as a positive mediator for angiogenesis. To investigate
the biologic function of AGF in angiogenesis, we analyzed the
vasculature in the dermis of transgenic mice expressing AGF
in mouse epidermal keratinocytes (K14-AGF). K14-AGF transgenic
mice were grossly red, especially in the ears and snout, suggesting
that hypervascularization had occurred in their skin. Histologic
examination of ear skin from K14-AGF transgenic mice revealed
increased numbers of microvessels in the dermis, whereas the
expression of several angiogenic factors, such as basic fibroblast
growth factor (bFGF), vascular endothelial growth factors (VEGFs),
and angiopoietin-1 (Ang-1), was decreased. We showed that AGF
is a secreted protein and does not bind to tyrosine kinase with
immunoglobulin and EGF-homology domain (Tie1) or Tie2 receptors.
An in vitro chamber assay revealed that AGF directly promotes
chemotactic activity of vascular endothelial cells. Both mouse
corneal and matrigel plug assays showed that AGF induces neovascularization
in vivo. Furthermore, we found that plasma leakage occurred
after direct injection of AGF into the mouse dermis, suggesting
that AGF directly induces a permeability change in the local
vasculature. On the basis of these observations, we propose
that AGF is a novel angiogenic factor and that handling of its
biologic functions could lead to novel therapeutic strategies
for control of angiogenesis.
Oike Y, et al. Blood. 2004 May 15;103(10):3760-5
Angiopoietin-related growth factor antagonizes obesity
and insulin resistance
Angiopoietin-related growth factor (AGF), a member of the angiopoietin-like
protein (Angptl) family, is secreted predominantly from the
liver into the systemic circulation. Here, we show that most
(>80%) of the AGF-deficient mice die at about embryonic day
13, whereas the surviving AGF-deficient mice develop marked
obesity, lipid accumulation in skeletal muscle and liver, and
insulin resistance accompanied by reduced energy expenditure
relative to controls. In parallel, mice with targeted activation
of AGF show leanness and increased insulin sensitivity resulting
from increased energy expenditure. They are also protected from
high-fat diet-induced obesity, insulin resistance and nonadipose
tissue steatosis. Hepatic overexpression of AGF by adenoviral
transduction, which leads to an approximately 2.5-fold increase
in serum AGF concentrations, results in a significant (P <
0.01) body weight loss and increases insulin sensitivity in
mice fed a high-fat diet. This study establishes AGF as a new
hepatocyte-derived circulating factor that counteracts obesity
and related insulin resistance.
Oike Y, et al. Nat Med. 2005 Apr;11(4):400-8. Epub 2005 Mar
20
|
|
|
|
|
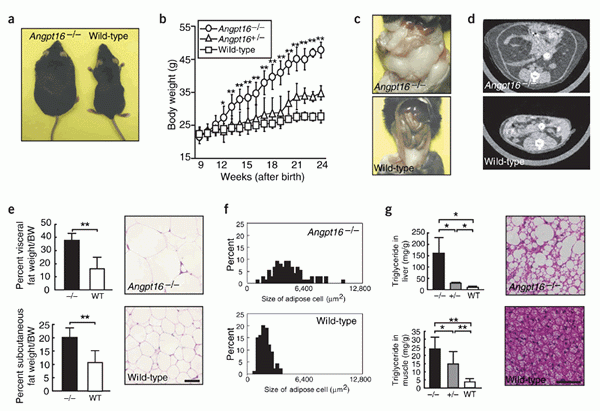
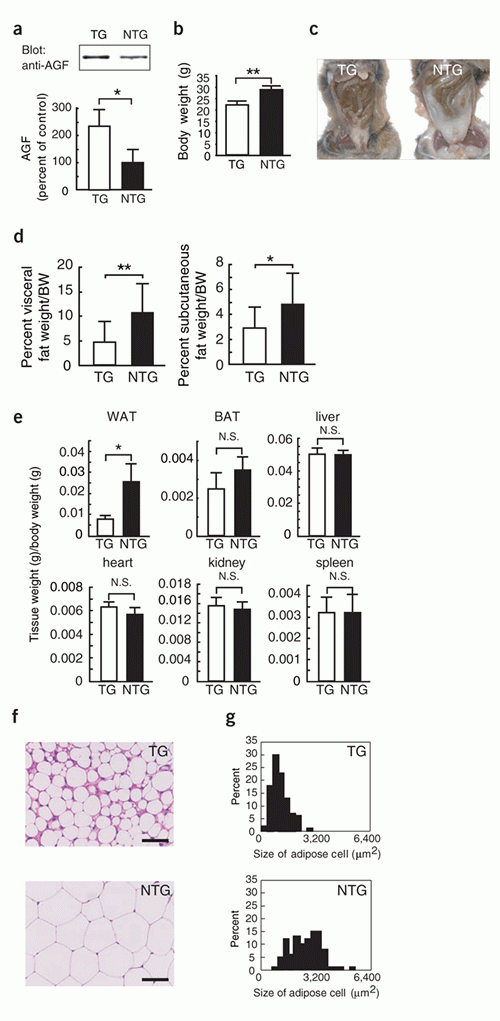
Obesity in Angptl6-/- mice on a normal diet.
(a) Gross appearance of Angptl6-/- mice and wild-type
control mice. (b) Body weight of each genotype (n = 8).
(c-g) Abdominal cavity (c), CT findings at a level of
8 mm above the top of the iliac bone (d), visceral fat
(n = 5) and subcutaneous fat (n = 5) weight/body weight,
and histological analysis (e) and distribution of cell
size (f) of WAT of Angptl6-/- mice and wild-type mice.
(g) Triglyceride levels in liver (n = 5) and gastrocnemius
muscle (n = 5), and hematoxylin and eosin-stained sections
of BAT of Angptl6-/- and wild-type mice. Data are mean
s.d. Bars in histological sections indicate 50 m. * P
< 0.05, ** P < 0.01, between the two genotypes indicated.
Female mice 8 months after birth were used for all experiments.
Oike Y, et al. Nat Med. 2005 Apr;11(4):400-8. Epub 2005
Mar 20 |
|
Obesity in Angptl6-/- mice on a normal diet.
(a) Gross appearance of Angptl6-/- mice and wild-type
control mice. (b) Body weight of each genotype (n = 8).
(c-g) Abdominal cavity (c), CT findings at a level of
8 mm above the top of the iliac bone (d), visceral fat
(n = 5) and subcutaneous fat (n = 5) weight/body weight,
and histological analysis (e) and distribution of cell
size (f) of WAT of Angptl6-/- mice and wild-type mice.
(g) Triglyceride levels in liver (n = 5) and gastrocnemius
muscle (n = 5), and hematoxylin and eosin-stained sections
of BAT of Angptl6-/- and wild-type mice. Data are mean
s.d. Bars in histological sections indicate 50 m. * P
< 0.05, ** P < 0.01, between the two genotypes indicated.
Female mice 8 months after birth were used for all experiments.
Oike Y, et al. Nat Med. 2005 Apr;11(4):400-8. Epub 2005
Mar 20 |
|
| (a) Western blotting analysis for serum AGF in Angptl6
transgenic (TG) and nontransgenic control (NTG) mice at
4 months of age. The ratio for the control is set as 100%.
(b-g) Body weight (b), and gross appearance of visceral
adipocyte (c) in TG and NTG mice at 4 months of age. (d)
Comparison of visceral fat and subcutaneous fat weight/body
weight between TG and NTG mice at 5 months of age. Tissue
weight/body weight in TG and NTG mice at 4 months of age
(e). n = 10-15 in each group. Histological analysis (f)
and distribution of cell size (g) of WAT from TG and NTG
mice at 4 months of age. Scale bars, 50 m. Data are mean
s.d. *P < 0.05, **P < 0.01, between the two genotypes
indicated. N.S. indicates no significant difference compared
with nontransgenic wild-type mice. Female mice were used
for all experiments. Oike Y, et al. Nat Med. 2005 Apr;11(4):400-8.
Epub 2005 Mar 20 |
|
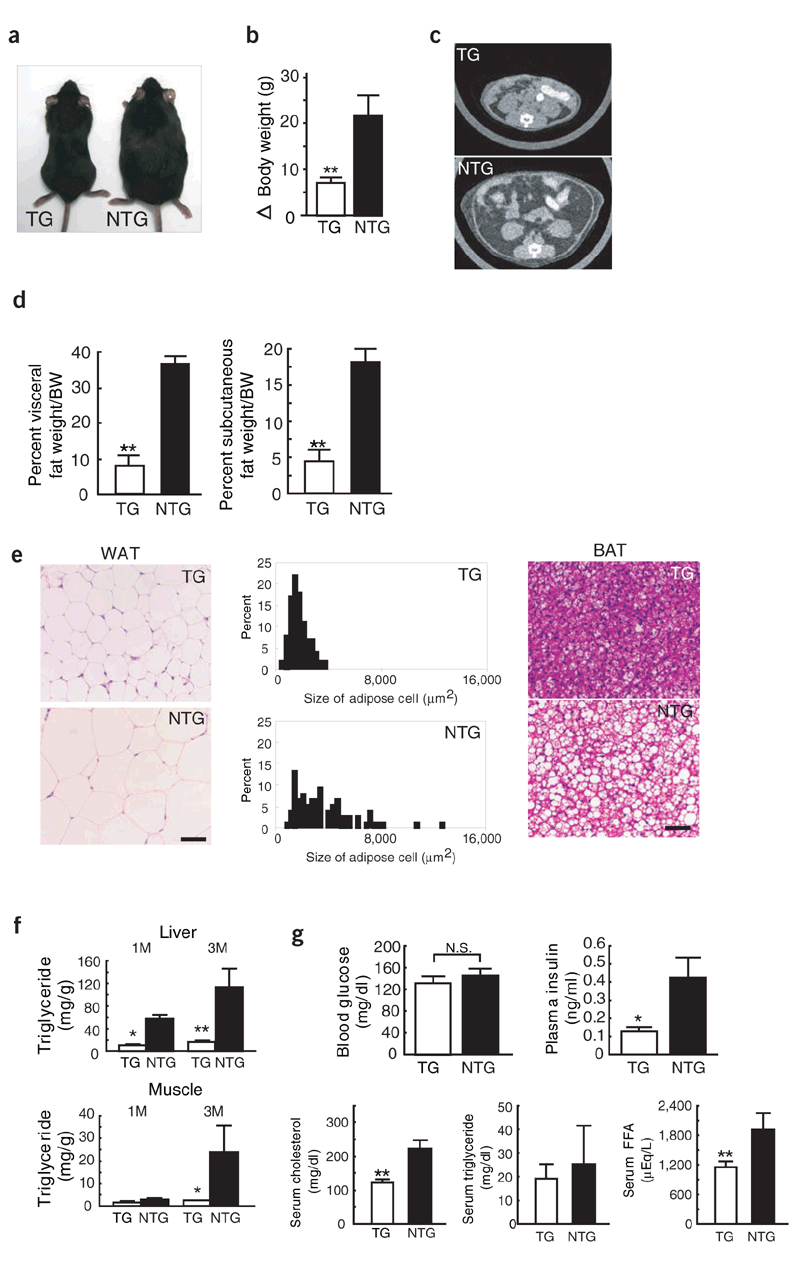
Resistance to high-fat diet-induced obesity and related
metabolic disorders seen in Angptl6 transgenic mice.
(a) Representative gross appearance of AGF-transgenic
(TG) and nontransgenic control (NTG) female mice after
3-months high-fat feeding. (b) Change in body weight in
TG and NTG male and female mice after 3-months high-fat
feeding (n = 10). (c) CT findings, which were shown at
a level of 8 mm above the top of the iliac bone in a.
(d) Visceral fat (n = 10) and subcutaneous fat (n = 10)
weight/body weight. (e) Hematoxylin and eosin-stained
sections and distribution of cell size of WAT, and hematoxylin
and eosin-stained sections of BAT from TG and NTG mice
after 3 months of high-fat feeding. (f) Triglyceride content
in liver (n = 5) and gastrocnemius muscle (n = 5) from
TG and NTG mice after 1 month (left) and 3 months (right)
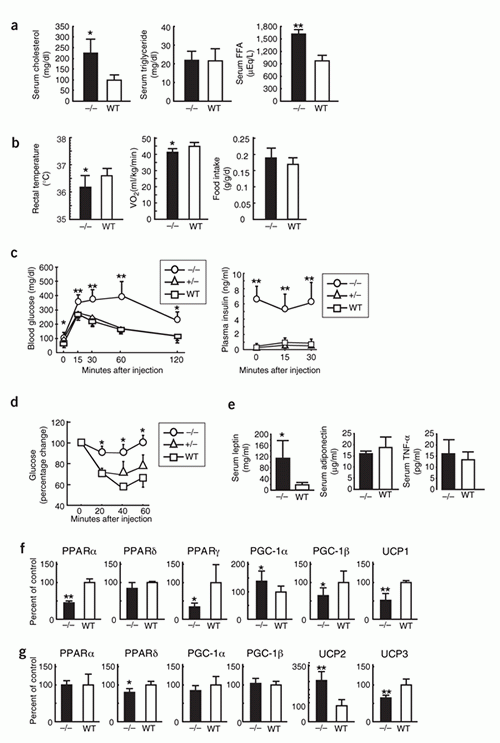
of high-fat feeding. (g) Blood glucose, plasma insulin,
serum cholesterol, triglyceride, and FFA concentrations
in TG and NTG mice after 3-months of high-fat feeding.
(n = 10 in each group). Scale bars in e, 50 m. Data are
mean s.d. * P < 0.05, ** P < 0.01, between the two
genotypes indicated.
|
|
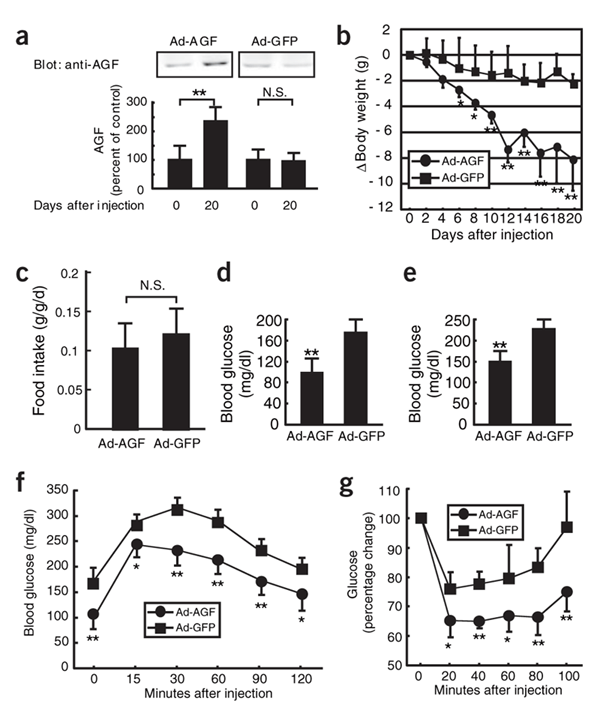
AGF decreased body weight and increased insulin sensitivity
in high-fat fed-induced obese mice.
(a) The relative ratio of serum concentrations of AGF
in Ad-AGF injected and Ad-GFP injected mice on day 20
relative to each mouse on day 0. The value of serum AGF
concentrations on day 0 is set at 100% (n = 5-8 in each
group). (b) Alteration in body weight of high-fat fed-induced
obese female mice after Ad-AGF and Ad-GFP injections (n
= 8 in each group). (c-g) Comparison of food intake/lean
body weight (c), fasting blood glucose (d), random fed
blood glucose (e), glucose tolerance test (f) and insulin
tolerance test (g) between Ad-AGF injected and Ad-GFP
injected mice (n = 5-8 in each group). Data are mean s.d.
* P < 0.05, ** P < 0.01, between the two groups.
N.S. indicates no significant difference compared with
Ad-GFP-injected mice. |
|
|
|
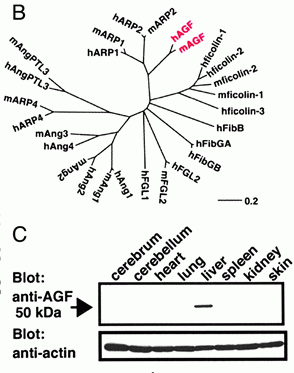
| Sequence and expression analyses of AGF. (A) Deduced
amino acid sequences of human and mouse AGF. Open and
filled arrows indicate the limits of the coiled-coil and
fibrinogen-like domains, respectively. (B) The evolutionary
relationship of AGF (red) to the angiopoietin superfamily
was derived by using DNASIS FOR WINDOWS V. 2.1 (Hitachi
Software, Tokyo). The length of each horizontal line is
proportional to the degree of amino acid sequence divergence.
(C) Western blot analysis of various mouse tissues by
using the anti-AGF (Upper) and anti-actin (Lower) antibodies.
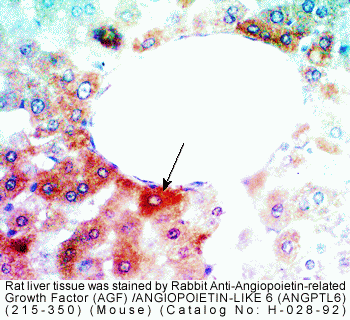
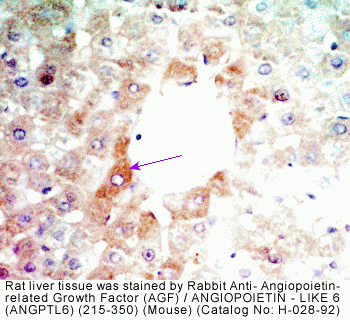
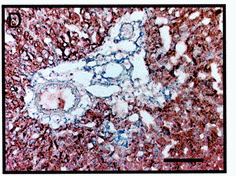
(D) Immunoreaction using the anti-AGF antibody shows AGF
specifically expressed in hepatic parenchymal cells, not
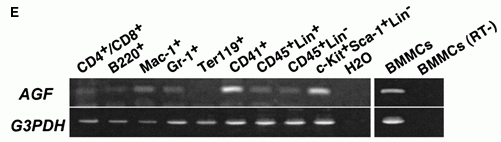
in the Glisson region. (Scale bar = 100 µm.) (E) Analysis
of AGF mRNA expression in hematopoietic cells from adult
bone marrow or BMMCs by RT-PCR. CD4+/CD8+, T cell; B220,
B cell; Mac-1, macrophage and monocyte; Gr-1, granulocyte;
Ter119, erythrocyte; CD41, megakaryocyte/platelet; CD45+Lin+,
mature HCs; CD45+Lin-, immature HCs; c-Kit+Sca-1+Lin-,
hematopoietic stem cell-enriched population. A mixture
of anti-Mac-1, -Gr-1, -B220, -CD4, -CD8, and -Ly-6 antibodies
was used as a lineage marker (Lin). GAPDH mRNA served
as a loading control. All RNA without RT treatment (RT-)
show no transcript by PCR. BMMCs (RT-) is one representative
data. Oike Y, et al. Proc Natl Acad Sci U S A. 2003 Aug
5;100(16):9494-9 |
|
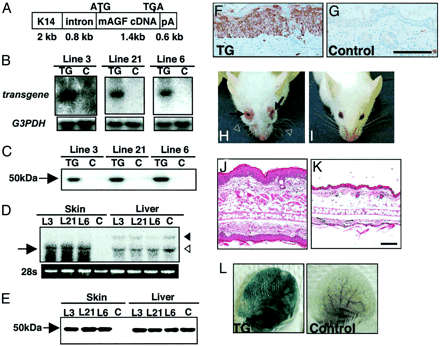
| Markedly thickened epidermal layers in K14-AGF mice.
(A) Schematic representation of the transgene used to
generate K14-AGF mice. K14, intron, and pA indicate the
human K14 promoter, rabbit -globin intron, and a polyadenylation
signal derived from the K14 gene, respectively. (B and
C) Expression of the transgene was detected in the whole
skin of F1 mice (TG) 3 days after birth by Northern (B)
and Western (C) blotting analysis. No expression of the
transgene was detected in controls (C). Blotting analysis
for GAPDH was performed as an internal control experiment.
(D and E) Comparison of the mRNA (D) and protein (E) level
of AGF from skin and liver between K14-AGF mice and controls.
Arrow in D indicates the transcription of the transgene.
Open and filled arrowheads in D indicate 1.8- and 4.0-kb
endogeneous AGF transcripts, respectively. Five micrograms
of protein was loaded in each lane in E.(F and G) Immunohistochemical
analysis of AGF detects expression of the transgene in
the epidermis of skin from the ears of F1 K14-AGF mice
(F) and their controls (G). (Scale bar = 100 µm.) (H and
I) Front view of the K14-AGF mouse and controls. Swelling
of the eyelid (arrows in H), ears, and nose, and wavy
whiskers (open arrowheads in H) were detected in K14-AGF
mice. (J and K) Hematoxylin/eosin histology of swollen
ear of the K14-AGF mouse (J) and control (K). (Scale bar
= 100 µm.) (L) Photographs of ears injected intravenously
with Evans blue dye to visualize plasma leakage. The ear
of a K14-AGF mouse was strongly blue, whereas the control
ear was not changed. One representative experiment is
shown. Oike Y, et al. Proc Natl Acad Sci U S A. 2003 Aug
5;100(16):9494-9 |
|
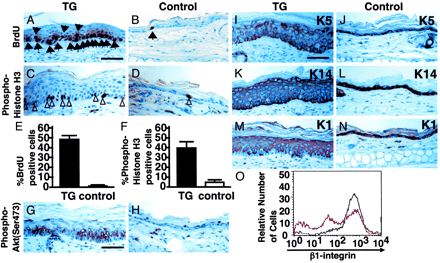
| Actively cycling expression of keratin proteins in epidermal
cells of K14-AGF mice. (A–F) Comparison of levels
of DNA synthesis in the epidermis from K14-AGF mice and
controls. Skin sections from both mice were stained immunohistochemically
for BrdUrd (A and B) and anti-phospho-histone H3 (C and
D)by using peroxidase-based detection. Sections were counterstained
with hematoxylin. Arrows and arrowheads indicate examples
(brown-stained nuclei) of BrdUrd-positive and phospho-histone
H3-positive cells, respectively. (E and F) The average
numbers of labeled cells with BrdUrd and anti-phospho-histone
H3 immunoreactivity from five sections each from three
mice, respectively. (G and H) Immunoreactivity against
anti-phospho-Akt antibody was seen in the thickened epidermis
from the K14-AGF mouse (G), whereas no immunoreactivity
was seen in epidermis from controls (H). (I–N)K5(J)
and K14 (L) were detected in the basal layer of the epidermis
in controls. Similar sections obtained from K14-AGF mice
show positive staining in the suprabasal layer of the
epidermis as well(I and K). K1 staining was similar for
both the K14-AGF mouse (M) and controls (N). (Scale bar
= 50 µm.) (O) Surface levels of 1-integrins in basal keratinocytes
of K14-AGF (red line) and controls (black line). Three
peaks for intensity of 1-integrin expression are detected
in K14-AGF, whereas one peak with high intensity is seen
in controls. Oike Y, et al. Proc Natl Acad Sci U S A.
2003 Aug 5;100(16):9494-9 |
|
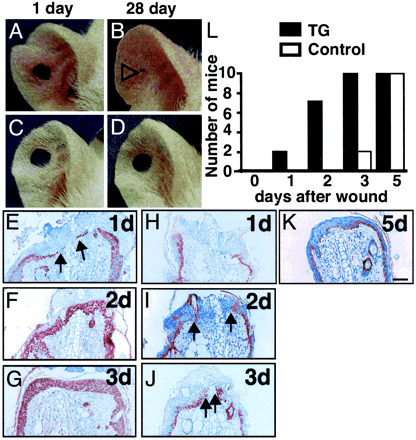
| Reepithelialization of wounds in K14-AGF mice. (A–D)
Representative photograph ear wound healing. K14-AGF and
control ears were punched in the center creating a 2-mm
open hole and followed for 28 days. (A and B) One can
see the progression of hole closure from day 1 (A) to
day 28 (B). Open arrowhead in B indicates shortened hole
in K14-AGF. (E–K) Wounding was accomplished by ear
segment excisions. Shown are representative data of skin
sections with staining for anti-K14 antibody from the
exposed portion of the remaining ear of K14-AGF mice (E–G)
and control littermates (H–K)at1,2,3, and 5 days
after the initial wounds. All sections were photographed
at the same magnification. (Scale bar = 100 µm.) Arrowheads
indicate migrating and proliferating epidermal keratinocytes,
indicating that keratinocytes overlapped the injury site
rapidly in K14-AGF mice. (L) Analysis of frequencies of
wound closure by histological examination at 1, 2, 3,
and 5 days after the initial wounding. Filled (controls)
and open (K14-AGF mice) columns represent the number of
mice in which the wound was completely covered with keratinocytes.
Ten mice were examined on each day of the experiment.
Oike Y, et al. Proc Natl Acad Sci U S A. 2003 Aug 5;100(16):9494-9 |
|
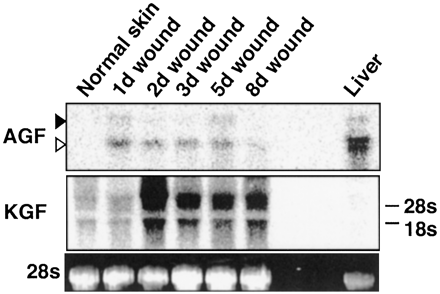
| Expression of AGF mRNA in normal and wounded skin.
The total RNA (10 µg) from normal and wounded ear skin
was analyzed by Northern blotting analysis with cDNA probes
for AGF and KGF. The relative amount of each mRNA was
quantified with normalization to 28S rRNA levels. The
time after injury is indicated on top of each lane: 1,
2, 3, 5, and 8 days. Oike Y, et al. Proc Natl Acad Sci
U S A. 2003 Aug 5;100(16):9494-9 |
|
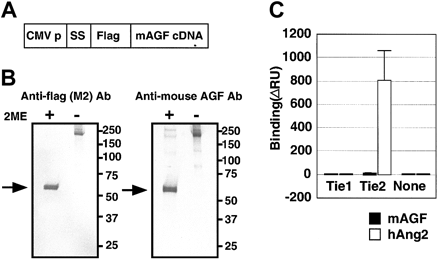
AGF and TIE receptors. AGF does not
bind to tyrosine kinase with immunoglobulin and EGF-homology
domain (Tie) receptors. (A) Schematic representation
of the plasmid construction used to generate FLAG-AGF
protein. CMV-p and SS indicate the cytomegalovirus promoter
and a signal sequence, respectively. (B) After transfection
of HEK293 cells with a mouse AGF cDNA with a 5'-terminal
extension encoding a FLAG-tag (panel A), mouse AGF-FLAG
fusion protein was detected in culture supernatants
by Western blot analysis with an anti-FLAG antibody
(left) and an antimouse AGF antibody (right), with or
without 2-mercaptoethanol (2-ME). Lanes contain approximately
10 ng purified protein. Arrows indicate the monomer
of FLAG-AGF protein. (C) Ties are not receptors for
AGF. BIAcore binding assay of AGF (200 ng) to the Tie1
and Tie2 receptors. As a positive control, human Ang-2-6xHis-tagged
protein (200 ng) specifically bound to the Tie2-Fc protein
(460 ng), but not to the Tie1-Fc (460 ng). Error bar
represents mean ± SD. Blood. 2004 May 15;103(10):3760-5.
Oike Y, et al. Blood. 2004 May 15;103(10):3760-5 |
|
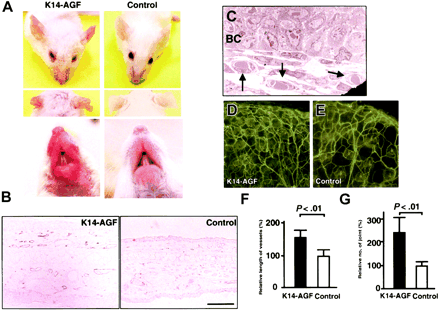
Increased number of microvessels in
K14-AGF transgenic mice. (A) Gross appearance of the
K14-AGF mouse and a control showing that the skin of
ears and snout of K14-AGF mice are red compared with
controls. (B) Immunohistochemical analysis with anti–PECAM-1
antibody of ear skin from K14-AGF mouse and a control.
Increased PECAM-1+ microvessels (purple) are detected
in the dermis and subcutaneous layers of K14-AGF mouse.
Bar indicates 100 µm. (C) Electron microscopic analysis
(original magnification, x 1700) shows that increased
vessels are capillary-sized (arrows) in the K14-AGF
mouse. BC indicates epidermal basal cells. (D-E) Representative
photograph of blood vessels in the ear from the K14-AGF
mouse (D) and controls (E) stained with fluorescein-labeled
Lycopersicon esculentum lectin. Abundant capillary-sized
vessels in the K14-AGF mouse are detected. (F-G) Quantitative
analysis for the number of vessels shown in panels D
and E. Length of vessels (F) and number of vessel joints
(G) in K14-AGF transgenic mice () relative to controls
() are shown as percentages. Columns represent mean
values + SD (n = 5). Oike Y, et al. Blood. 2004 May
15;103(10):3760-5 |
|