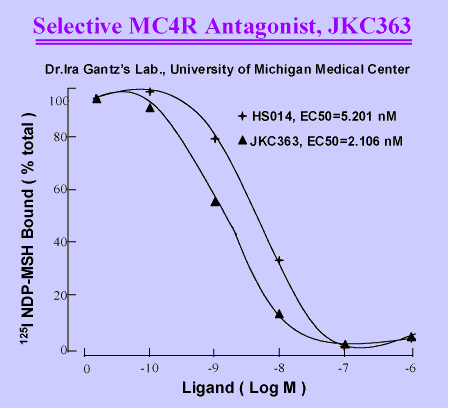
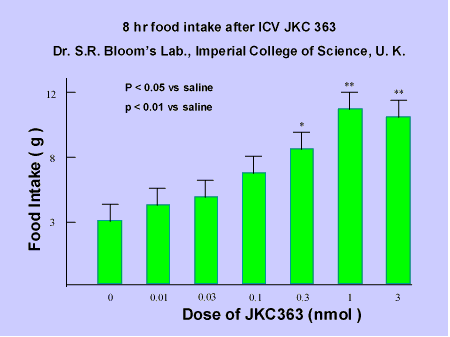
Compound 7 An highly
selective antagonist for MC3R
more
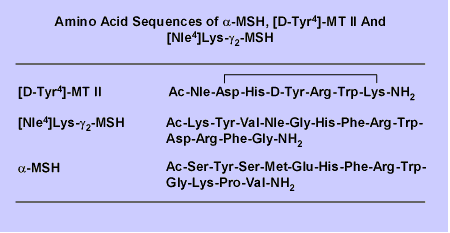
information for Alpha MSH, Gramma-MSH, Beta-MSH, AGRP,
VA-beta-MSH, MT II, SHU9119
Structure-activity
relationships of novel cyclic alpha-MSH/beta-MSH hybrid
analogues that lead to potent and selective ligands
for the human MC3R and human MC5R.
It
has been shown by extensive studies that alpha-MSH
bioactivity is critically dependent on the core or
central tetrapeptide sequence, His-Phe-Arg-Trp, however
with poor selectivity for the human MC3R-MC5R. The
structure-activity relationships study here is aimed
at identifying lead structures or templates of this
core sequence by the use of different conformational
constraints that might impart changes in its topography
and thus promote differences in potency and selectivity
at these receptors. Our peptide library consists of
a novel series of cyclic alpha-MSH analogues that
have disulfide bridges between Cys or Cys-like residues
at positions 4 and 10, giving rise to 23-membered
rings fused at the C-terminal end with the C-terminal
fragment of beta-MSH (Pro-Pro-Lys-Asp). While such
constraints of the peptide backbone with disulfide
bridges of different chirality affect potency and
selectivity at these receptors, further changes in
the hydrophobicity at position 7 with either a D-Phe
or D-Nal(2') and replacement of a His with a Pro in
position 6 cause additional effects. Thus, the most
interesting lead compounds that emerged from this
study are (1) compound 5, Ac-c[Cys-Glu-His-D-Phe-Arg-Trp-D-Cys]-Pro-Pro-Lys-Asp-NH(2)
(IC(50) = 10 nM), which is the first potent and highly
selective antagonist ligand for the hMC5R (560-fold
vs the MC3R and 1000-fold vs the MC4R); (2) compound
7, Ac-c[Cys-Glu-Pro-D-Nal(2')-Arg-Trp-Cys]-Pro-Pro-Lys-Asp-NH(2)
(IC(50) = 31 nM), which is a highly selective antagonist
analogue for the MC3R (560-fold vs the hMC4R and about
3000-fold vs the hMC5R; and (3) compound 9, Ac-c[Pen-Glu-His-D-Nal(2')-Arg-Trp-Cys]-Pro-Pro-Lys-Asp-NH(2)
(IC(50) = 3 nM), which is more potent than 7 at the
MC3R but not as selective.
Balse-Srinivasan
P, Grieco P, Cai M, Trivedi D, Hruby VJ. J Med Chem.
2003 Aug 14;46(17):3728-33
Ligand
MC3R(IC50)
MC3R/MC4R
MC3R/MC5R
Compound
7
31 nM
560 fold
3000 fold
We report the isolation of a novel human circulating
proopiomelanocortin-derived peptide named VA-ß-MSH
from hemofiltrate and its pharmacological characterization.
Screening for lipolytic activity in differentiated
3T3-L1 adipocytes led to the isolation from a hemofiltrate
peptide library by alternating reversed-phase and
cation-exchange chromatography. In the course of this
isolation, we also identified human ß-MSH (1-22).
We synthesized VA-ß-MSH by the Fmoc (N-(9-fluorenyl)-methoxycarbonyl)
solid phase method and used syntheticß-MSH (1-22)
to confirm that both isolated peptides are lipolytically
active in a dosedependent manner in differentiated
3T3-L1 adipocytes in the nanomolar range. Using cAMPELISA,
we demonstrate that stimulation with both peptides
caused a strong cAMP elevation in this cell system.
Furthermore, we show that the selective inhibitors
of cAMP-dependent protein kinase, Rp-8-CPT-cAMPS and
H89, significantly reduce VA-ß-MSH- andß-MSH (1-22)-mediated
lipolysis. Although isolated here following its lipolytic
activity on 3T3-L1 cells, this newly identified circulating
human melanocortin may also serve other functions
in human physiology. Moreover, the fact that these
peptides have been identified following a functional
assay but have been overseen in large proteomic approaches
underscore the importance for such approaches in order
to identify previously undescribed circulating bioactive
molecules.
Isolation
and characterisation of a novel POMC-derived peptide
from hemofiltrate of chronic renal failure patients.
Fricke K., et al. Endocrinology.
First published January 13, 2005 as doi:10.1210/en.2004-1097
Lipolytic
potency and intracellular cAMP formation of VA-ß-MSH,
ß-MSH (1-22), hACTH (1-39), and forskolin. Results
are shown as means (± SD) of three independent experiments.
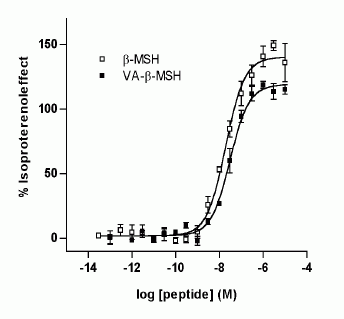
Lipolytic effect of
synthetic VA-ß-MSH and ß-MSH (1-22). Dose-response
relationships of increasing concentrations
of both peptides. After incubation of 3T3-L1
adipocytes with either peptide for 5 h,
glycerol content in the supernatants was
measured. Results are expressed as percentages
of the lipolytic response of cells to 1
µM isoproterenol and are means ± SD (n=3).
One of three independent experiments is
presented.
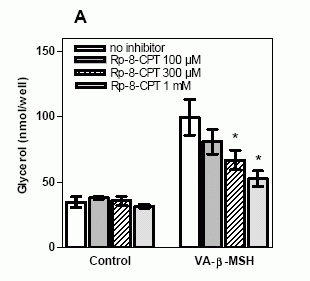
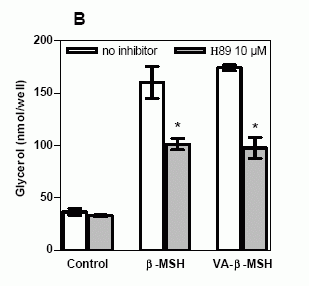
Effects
of specific PKA inhibitors on VA-ß-MSH-
and ß-MSH (1-22)- induced lipolysis. 3T3-L1
adipocytes were preincubated with or without
inhibitor for 40 minutes, followed by the
incubation with 20 nM VA-ß-MSH or ß-MSH
(1-22) for 5 h. A. Preincubation of 3T3-L1
adipocytes with 0.3 mM and 1 mM Rp-8-CPT-cAMPS
significantly decreased VA-ß-MSH-induced
lipolysis. B. VA-ß-MSH- and ß-MSH (1-22)-
induced glycerol release were inhibited
in the presence of H89. Basal lipolysis
was not significantly changed by either
inhibitor. Data points are means ± SD (n=3).
Results shown are representative for three
separate experiments. * P < 0.05 versus
no inhibitor.
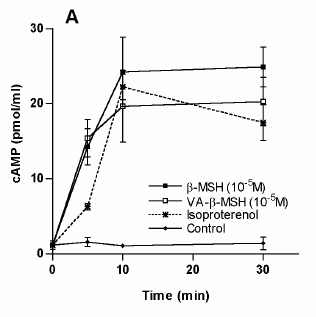
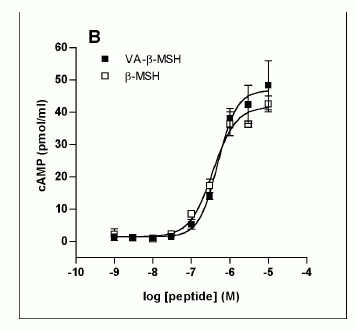
Characterization
of the VA-ß-MSH- and ß-MSH (1-22)- stimulated
cAMP production in 3T3-L1 adipocytes. The
differentiated adipocytes were transferred
into fresh DMEM on the day of the experiment,
varying concentrations of ß-MSH (1-22) and
VA-ß- MSH were added and incubated at 37°C
for the indicated time points. The supernatants
were 27 removed and cells were lysed with
seventy percent ethanol. Cell lysates were
assayed for cAMP by ELISA. A. Time dependency
of VA-ß-MSH- and ß-MSH (1-22)- stimulated
cAMP production in 3T3-L1 cells. Isoproterenol
(1 µM) was used as a positive control. B
Doseresponse relationship of cAMP-production
in differentiated 3T3-L1 cells by VA-ß-MSH
andß-MSH (1-22). Each point represents the
mean ± SD of triplicate values. Results
are representative for three (A) and two
(B) independent experiments.
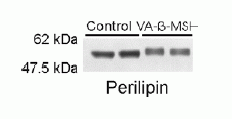
Effect
of VA-ß-MSH on CREB and perilipin phosphorylation.
A 3T3-L1 adipocytes were serum deprived
overnight and subsequently stimulated with
300 nM VA-ß- MSH for the times indicated.
At each time point, cells were lysed, and
equal amounts of lysate protein were subjected
to immunoblot analysis. Blots were probed
with antibodies against serine 133-phosphorylated
CREB (pCREB) and total CREB. One of three
independent experiments each with two replicates
is shown. The positions of the pCREB and
CREB bands are indicated. B After 15 min
stimulation with VA-ß-MSH, cell lysates
were subjected to immunoblot analysis, and
blots were probed with an antibody against
perilipin. The characteristic upward shift
observed with VA-ß-MSH reveals the phosphorylation
of perilipin A in stimulated cells compared
to cells treated with control medium (representative
blot of three independent experiments).
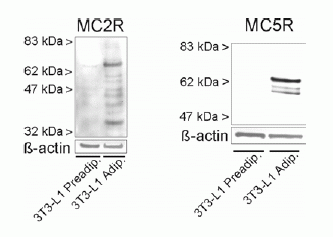
Protein expression
of melanocortin receptor isoforms 2 and
5 in 3T3-L1 adipocytes. Untreated 3T3-L1
adipocytes were lysed and subjected to SDS-PAGE
followed by immunoblot analysis. Blots were
probed using a specific antiserum directed
against MC2R (left side) or an affinity-purified
antibody against MC5R (right side). ß-actin
served as a loading control.
Fricke
K., et al. Endocrinology. First published January 13,
2005 as doi:10.1210/en.2004-1097
beta-MSH: a functional ligand that regulated energy
homeostasis via hypothalamic MC4-R?
alpha-Melanocyte stimulating hormone (MSH) has generally
been assumed to be the endogenous ligand acting at
the melanocortin-4 receptor (MC4-R), activation of
which in the hypothalamus leads to reduced feeding.
However, beta-MSH is also capable of activating MC4-R
and inhibiting feeding. Here, we investigated the
possibility that beta-MSH acts as an endogenous MC4-R
agonist and that this melanocortin peptide plays a
role in the regulation of feeding and energy balance.
We found that beta-MSH had significantly higher affinities
than alpha-MSH at both human MC4-R transfected into
CHO cells (K(i): beta-MSH, 11.40.4 nmol/l versus alpha-MSH,
32416 nmol/l, P<0.001) and MC4-R in rat hypothalamic
homogenates (K(i): beta-MSH, 5.00.4 nmol/l versus
alpha-MSH, 22.52.3 nmol/l, P<0.001). Incubation
of brain slices with 5 microM beta-MSH significantly
increased [35S]GTPgammaS binding by 140-160% (P<0.001),
indicating activation of G-protein-coupled receptors
(GPCRs), in the hypothalamic ventromedial (VMH), dorsomedial
(DMH), arcuate (ARC) and paraventricular (PVN) nuclei.
These sites match the distribution of beta-MSH immunoreactive
fibres and also the distribution of MC4-R binding
sites which we and others previously reported. Food-restriction
significantly increased beta-MSH levels in the VMH,
DMH and ARC (all P<0.05) above freely-fed controls,
whilst alpha-MSH concentrations were unchanged. We
propose that increased beta-MSH concentrations reflect
blockade of the peptide's release in these sites,
consistent with the increased hunger and the known
up-regulation of MC4-R in the same nuclei. Thus, we
conclude that (1). beta-MSH has higher affinity at
MC4-R than alpha-MSH; (2). beta-MSH activates GPCR
in these sites, which are rich in MC4-R; and (3).
beta-MSH is present in hypothalamic nuclei that regulate
feeding and its concentrations alter with nutritional
state. We suggest that beta-MSH rather than alpha-MSH
is the key ligand at the MC4-R populations that regulate
feeding, and that inhibition of tonic release of beta-MSH
is one mechanism contributing to hunger in under-feeding.
Harrold JA, et al. Peptides.
2003 Mar;24(3):397-405
Binding
Assay (IC50, nM)
Selectivity
hMC-3R
hMC-4R
hMC-5R
3/4
5/4
MC-4R Agonist
490+-5
4.3+-.67
4600+-50
114
1070
MT II
1.6+-.09
0.07+-.02
0.89+-.01
23
13
Melanocyte
Stimulating Hormones (MSH) Related Products