|
|
|
LGR7 (Relaxin Receptor 1) and LGR8 (Relaxin Receptor 2) |
Two Newly Identified orphan G Protein-Coupled Receptors for Insulin-like peptide 3 & 7 |
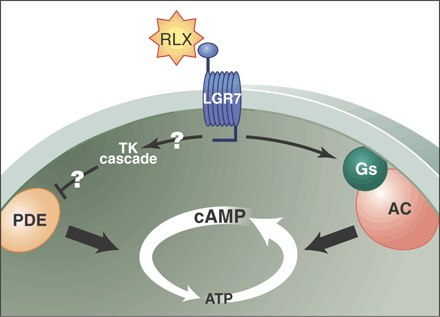
The different signal transduction pathways involved in the
up-regulation of cAMP by the peptide hormone relaxin (RLX). When
relaxin binds to its G protein-coupled receptor, LGR7 or LGR8, a G
protein signaling pathway is activated leading to stimulation of
adenylate cyclase (AC) and an increase in cAMP. Binding of relaxin
to its receptor also may activate a tyrosine kinase pathway that
inhibits the activity of a phosphodiesterase (PDE) that degrades
cAMP .
Richard Ivell. Science, Vol 295, Issue 5555, 637-638 , 25 January 2002
| |
LGR7 |
LGR8 |
|
Max* |
IC50 |
Max |
IC50 |
| INSL3 |
442.4 ± 2.1 |
32.9 ± 9.1 |
ND^ |
1576 ± 592 |
| INSL7/H3 Relaxin |
ND |
ND |
336.9 ± 12.3 |
0.19 ± 0.08 |
| |
IC50 values are in nM |
|
*Max, maximal levels of cAMP production
(pmol/2 ?105 cells) |
| ^ND, not determined due to
nonsaturation |
|
Sudo S, et al. J Biol Chem. 2003 Mar
7;278(10):7855-62. Epub 2002 Dec 27. |
Leucine-rich repeat-containing, G protein-coupled receptors (LGRs) represent a unique subgroup of G protein-coupled receptors with a large ectodomain. Recent studies demonstrated that relaxin activates two orphan LGRs, LGR7 and LGR8, whereas INSL3/Leydig insulin-like peptide specifically activates LGR8. Human relaxin 3 (H3 relaxin) was recently discovered as a novel ligand for relaxin receptors. Here, we demonstrate that H3 relaxin activates LGR7 but not LGR8. Taking advantage of the overlapping specificity of these three ligands for the two related LGRs, chimeric receptors were generated to elucidate the mechanism of ligand activation of LGR7. Chimeric receptor LGR7/8 with the ectodomain from LGR7 but the transmembrane region from LGR8 maintains responsiveness to relaxin but was less responsive to H3 relaxin based on ligand stimulation of cAMP production. The decreased ligand signaling was accompanied by decreases in the ability of H3 relaxin to compete for (33)P-relaxin binding to the chimeric receptor. However, replacement of the exoloop 2, but not exoloop 1 or 3, of LGR7 to the chimeric LGR7/8 restored ligand binding and receptor-mediated cAMP production. These results suggested that activation of LGR7 by H3 relaxin involves specific binding of the ligand to both the ectodomain and the exoloop 2, thus providing a model with which to understand the molecular basis of ligand signaling for this unique subgroup of G protein-coupled receptors.
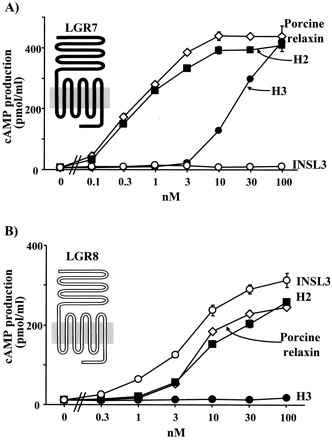
LGR7 is activated by porcine relaxin and H2 relaxin as well as
H3 relaxin, whereas LGR8 is activated by porcine relaxin, H2
relaxin, and INSL3. A, ligand-stimulated cAMP production mediated by
LGR7. B, ligand-mediated cAMP production mediated by LGR8. Purified
porcine relaxin, recombinant H2 relaxin, synthetic H3 relaxin, and
synthetic INSL3 were used.
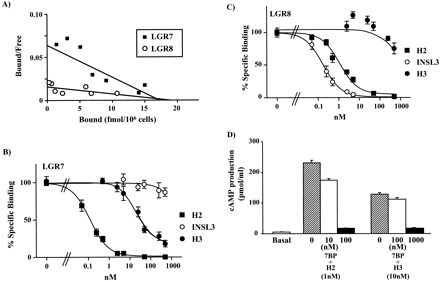
Direct binding of 33P-labeled H2 relaxin to LGR7 and LGR8 and
competition by relaxin-related peptides. A, Scatchard plot analyses
of 33P-labeled H2 binding to LGR7 and LGR8. B, competition of
33P-labeled H2 relaxin binding to LGR7 by relaxin-related peptides.
C, competition of 33P-labeled H2 relaxin binding to LGR8 by
relaxin-related peptides. D, the soluble ectodomain of LGR7 (7BP)
blocks H3 relaxin stimulation of cAMP production by LGR7-expressing
cells.
Sudo S, et al. J Biol Chem. 2003 Mar 7;278(10):7855-62. Epub
2002 Dec 27.
Relaxin is a hormone important for the growth and remodeling of reproductive and other tissues during pregnancy. Although binding sites for relaxin are widely distributed, the nature of its receptor has been elusive. Here, we demonstrate that two orphan heterotrimeric guanine nucleotide binding protein (G protein)-coupled receptors, LGR7 and LGR8, are capable of mediating the action of relaxin through an adenosine 3',5'-monophosphate (cAMP)-dependent pathway distinct from that of the structurally related insulin and insulin-like growth factor family ligand. Treatment of antepartum mice with the soluble ligand-binding region of LGR7 caused parturition delay. The wide and divergent distribution of the two relaxin receptors implicates their roles in reproductive, brain, renal, cardiovascular, and other functions.
Hsu S.Y., et al. Science 2002 Jan 15;295(5555):671-4
During male development testes descend from their embryonic intraabdominal position into the scrotum. Two genes, encoding the insulin-like 3 peptide (INSL3) and the GREAT/LGR8 G protein-coupled receptor, control the differentiation of gubernaculum, the caudal genitoinguinal ligament critical for testicular descent. It was established that the INSL3 peptide activates GREAT/LGR8 receptor in vitro. Mutations of Insl3 or Great cause cryptorchidism (undescended testes) in mice. Overexpression of the transgenic Insl3 causes male-like gubernaculum differentiation, ovarian descent into lower abdominal position, and reduced fertility in females. To address the question whether Great deletion complements the mutant female phenotype caused by the Insl3 overexpression, we have produced Insl3 transgenic mice deficient for Great. Such females had a wild-type phenotype, demonstrating that Great was the only cognate receptor for Insl3 in vivo. We have established that pancreatic HIT cells, transfected with the INSL3 cDNA, produce functionally active peptide. Analysis of five INSL3 mutant variants detected in cryptorchid patients showed that P49S substitution renders functionally compromised peptide. Therefore, mutations in INSL3 might contribute to the etiology of cryptorchidism. We have also showed that synthetic insulin-like peptides (INSL4 and INSL6) were unable to activate LGR7 or GREAT/LGR8.
Bogatcheva NV, et al. Mol Endocrinol. 2003 Dec;17(12):2639-46. Epub 2003 Aug 21.
 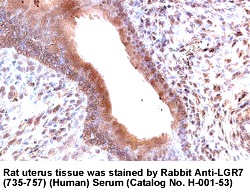
|
LGR7 (735-757), (Human) Antibody for
IHC |
|
Tissue
Sample
|
Rat
uterus
|
|
Fixative
|
10%
formalin
|
|
Embedding
|
paraffin
|
|
Negative Control
|
No
primary antibody
|
|
Pretreatment
|
Intact
|
|
Blocking
|
2%
Normal Goat Serum, 3%H2O2
|
|
Primary Antibody
|
LGR7 (735-757), (Human) Antiserum (Catalog No.:
H-001-53)
|
|
Optimal Dilution
|
1:100
(1
hour at RT )
|
|
Secondary Antibody
|
Goat
Anti-Rabbit IgG, Biotinylated (1:400)
|
|
Amplification
|
ABC
(Vector)
|
|
Detection System
|
HRP
|
|
Substrate
|
DAB
(Sigma)
|
|
Counterstained
|
Hematoxylin
|
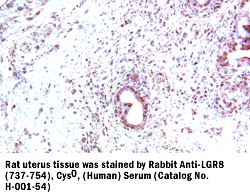 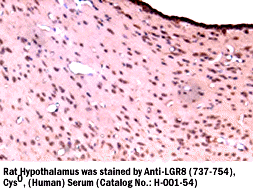
|
LGR8(737-754),
Cys0, (H) Antibody for IHC |
|
Tissue Sample
|
Rat hypothalamus |
|
Fixative
|
10% formalin
|
|
Embedding
|
paraffin
|
|
Negative Control
|
No primary antibody
|
|
Pretreatment
|
Pronase
|
|
Blocking
|
2% Normal Goat Serum, 3%H2O2
|
|
Primary Antibody
|
LGR8 (737-754), Cys0, (Human)
Antiserum (Catalog No.:
H-001-54)
|
|
Optimal Dilution
|
1:100
(1 hour at RT
)
|
|
Secondary Antibody
|
Goat Anti-Rabbit IgG, Biotinylated (1:400)
|
|
Amplification
|
ABC (Vector)
|
|
Detection System
|
HRP
|
|
Substrate
|
DAB (Sigma)
|
|
Counterstained
|
Hematoxylin
|
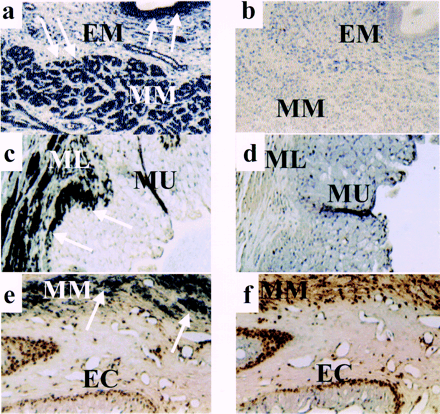
Tissue distribution of LGR7, as assessed by
immunohistochemical analysis in rodent tissues. Uterine
tissues were obtained from postpartum rats; vagina and cervix
were from rats at 19 days of pregnancy. Specific staining with
antibody to LGR7 (14) in uterus (a), vagina (c), and cervix
(e) is indicated by arrows. Nonimmune serum showed negligible
staining in uterus (b), vagina (d), and cervix (f).
Abbreviations: EM, endometrium; MM, myometrium; ML, muscularis
layer; MU, mucosal layer; EC, endocervix. Immunohistochemical
analysis was performed as described (6). Magnifications, ?00.
Hsu S.Y., et al. Science 2002 Jan 15;295(5555):671-4
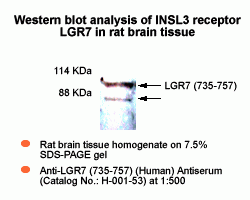
LGR7 antibody
is available for western blot analysis
 
|
|
|
Relaxin;Relaxin_E;Relaxin-2
%001-53%;%001-54%
|
|
|



