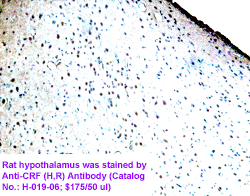
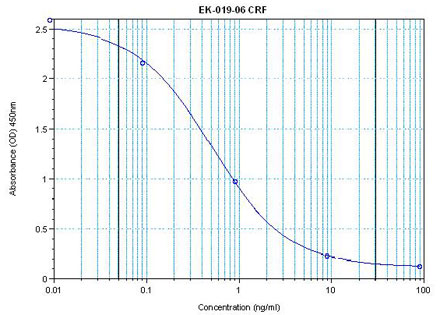
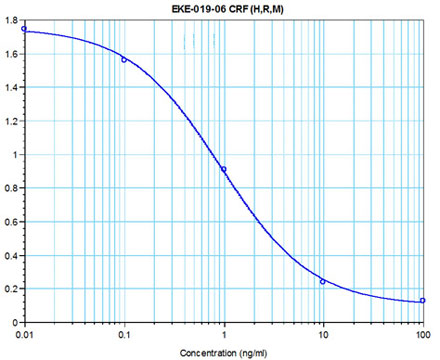
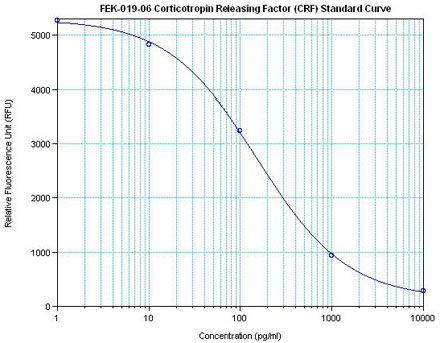
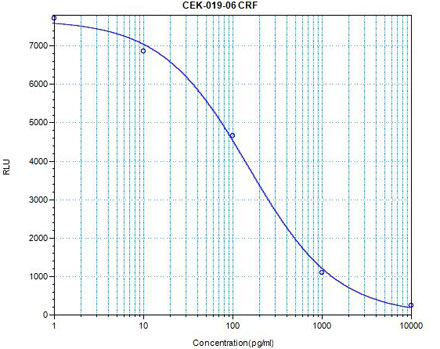
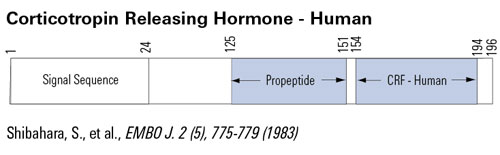
The view that energy balance is regulated has gained
acceptance in recent years. An important role in this
regulation is played by brain circuitries involved in the
control of energy intake (food intake) and energy expenditure
(thermogenesis) that are capable of integrating peripheral
signals, produced by perturbations of adipose tissue mass,
into messages to effectors of food intake and energy
expenditure, so as to prevent substantial variations in the
level of energy reserves. More than one neurosystem has been
reported to genuinely participate in the regulation of energy
balance. Among them is the corticotropin-releasing hormone
(CRH) system. This system, with its numerous clusters of brain
neurons, its closely related peptide urocortin, its two
receptor types and its binding protein, all generally widely
distributed throughout the brain, forms a network of neuronal
pathways capable of interacting with the circuitries
controlling food intake and energy expenditure. In addition,
CRH and urocortin's anorectic and thermogenic actions appear
to be coordinated to optimize energy losses. Finally, the CRH
system seems to demonstrate a certain degree of plasticity in
obesity and in response to food deprivation that is consistent
with its action on food intake and thermogenesis. The
observations have been made that food deprivation and obesity
can blunt the expression of the CRH type 2alpha receptor in
the ventromedial hypothalamic nucleus and can induce the
expression of the CRH-binding protein (a CRH-inactivating
protein) in brain areas involved in the anorectic and
thermogenic actions of CRH.
Richard D, Huang Q, Timofeeva E. The
corticotropin-releasing hormone system in the regulation of
energy balance in obesity. Int J Obes Relat Metab Disord 2000
Jun;24 Suppl 2:S36-9
The corticotropin-releasing factor (CRF) system could play
a significant role in the regulation of energy balance. This
system, which includes CRF, CRF-related peptides and CRF
receptors, is part of a huge network of cells connected to
central and peripheral pathways modulating energy metabolism.
CRF and CRF-related peptides, which elicit their effects
through G-protein-coupled receptors known in mammals as CRF(1)
receptor and CRF(2) receptor, are capable of strong anorectic
and thermogenic effects. Also supporting a role for the CRF
system in the regulation of energy balance are findings
demonstrating alterations in this system in obese and
food-deprived animals that concur to facilitate energy
deposition. In recent years, great progress has been made in
understanding the specific physiological roles of the CRF
system. In that respect, the discovery of urocortins II and
III, two endogenous ligands of the CRF(2) receptor, and the
development of selective and long-acting antagonists for the
CRF receptors, have led to a better comprehension of the role
of the CRF system in the regulation of energy balance.
Although there are still important unresolved issues in the
field of CRF research, the progress made recently warrants
investigations aimed at evaluating the CRF system as a
potential target for anti-obesity drugs.
Richard D, Lin Q, Timofeeva E. The
corticotropin-releasing factor family of peptides and CRF
receptors: their roles in the regulation of energy balance.
Eur J Pharmacol 2002 Apr
12;440(2-3):189-97
Corticotropin-releasing factor (CRF), urocortin and
urotensin I share amino acid sequences, and they inhibit food
intake in mammals. CRF plays a potent role in decreasing food
intake in avian species, but the effects of urocortin and
urotensin I have not been investigated. Therefore, the effect
of these three peptides on food intake in the neonatal chick
was compared. In Experiment 1, birds were injected
intracerebroventricularly (i.c.v.) with either 0, 0.01, 0.1 or
1 microg of urocortin following a 3-h fast, and food intake
was measured for 2 h post-injection. Food intake was
suppressed in a dose-dependent manner. Using a similar design
in Experiment 2, the effect of urotensin I was investigated.
Urotensin I appeared to suppress food intake in neonatal
chicks more than urocortin did. In Experiment 3, the efficacy
of CRF, urocortin and urotensin I was directly compared using
one dose, 0.1 microg. The results indicated that the
suppressive effect on food intake was strongest for CRF
followed by urotensin I, then urocortin. These results suggest
that the structure of receptors for the CRF family in chicks
may be somewhat different than in mammals.
Zhang R, Nakanishi T, Ohgushi A, Ando
R, Yoshimatsu T, Denbow DM, Furuse M. Suppression
of food intake induced by corticotropin-releasing factor
family in neonatal chicks. Eur J Pharmacol 2001 Sep
7;427(1):37-41
CRF-Receptor 2 / CRH-R2
A G-Protein-Coupled Receptor

Schematic representation of amino acids within the CRH/CRH-related agonists sequence important for determining CRH-R subtype selectivity. It is now accepted that amino acid residues 32–41 are important for receptor binding, whereas residues 1–16 are responsible for both binding and receptor activation. Residues present in the domain 17–31 appear to function as a linker providing the appropriate spatial and conformational support for the two binding regions. CRH-R2 selective agonists contain a proline at position 11 and alanine residues at positions 35 and 39 (the numbering of residues is based on h/rCRH sequence). In contrast, CRH-R nonselective peptides contain an arginine at position 35 and an acidic amino acid at position 39.
Edward W. Hillhouse and Dimitris K. Grammatopoulos . Endocrine Reviews 27 (3): 260-286