|
|
|
Cholecystokinin |
No cross-reactivity with Gastrin |
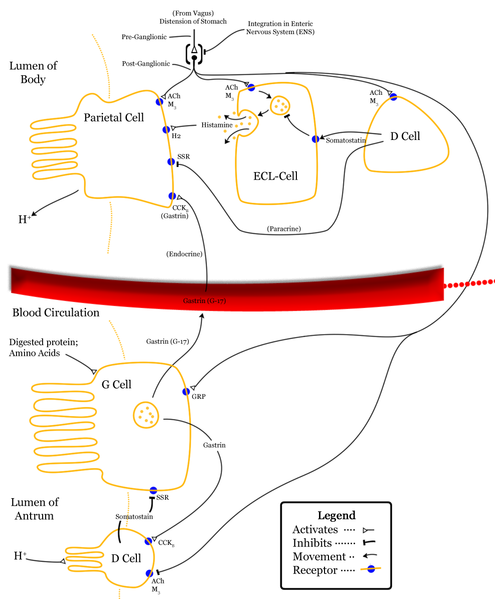
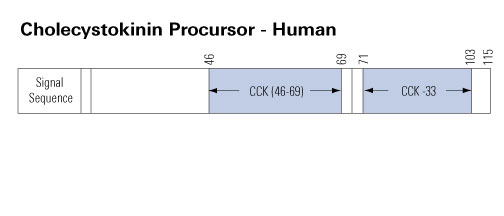
Cholecystokinin is a peptide hormone of the gastrointestinal system responsible for stimulating the digestion of fat and protein. Cholecystokinin, previously called pancreozymin, is synthesised by I-cells in the mucosal epithelium of the small intestine and secreted in the duodenum, the first segment of the small intestine, and causes the release of digestive enzymes and bile from the pancreas and gallbladder, respectively. It also acts as a hunger suppressant. Recent evidence has suggested that it also plays a major role in inducing drug tolerance to opioids like morphine and heroin, and is partly implicated in experiences of pain hypersensitivity during opioid withdrawal.
Hindbrain Response Model
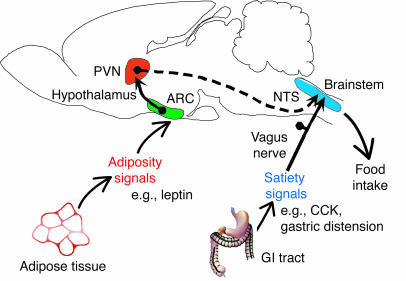
Model for regulation of the hindbrain response to satiety signals by hormonal input from the ARC. Adiposity signals such as insulin and
leptin circulate in proportion to body fat mass and act on hypothalamic ARC neurons
that project to hypothalamic areas such as the LHA (not shown) and PVN. In turn,
these “second order” neurons project to hindbrain autonomic centers
such as the NTS that process afferent input from satiety signals such as CCK. Input from descending, leptin-sensitive hypothalamic projections is integrated in the NTS with vagally mediated input from CCK, such that the timing of meal termination is regulated by changes in body fat content. Modified with permission from Nature GI, gastrointestinal.
Morton G.J., et al. J Clin Invest. 2005 March 1; 115(3):703–710.
Neurocentric model
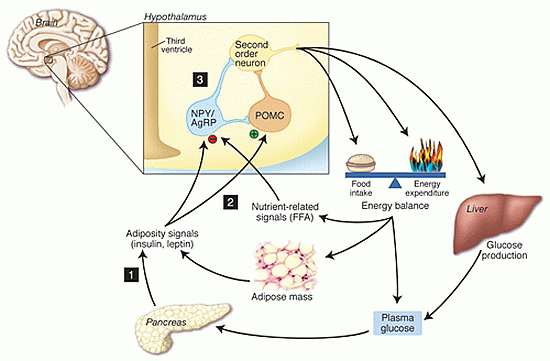
Neurocentric model depicting sites where defects in the negative feedback regulation of energy balance and glucose production predispose to weight gain and insulin resistance. Defects in the secretion of insulin or leptin (1), in the hypothalamic sensing of adiposity- or nutrient related signals (2), or in the neuronal responsiveness to these inputs (3) predispose to both positive energy balance and increased glucose production. If sustained, these will result in pathological weight gain and insulin resistance.
Schwartz MW, Porte D Jr. Diabetes, obesity, and the brain. Science. 2005 Jan 21;307(5708):375-9
We investigated the changes in the capacity for synthesis
of the exocrine pancreas of piglets during the 2 wk after weaning at 7 d of age
(trial 1) by measuring the expression of digestive enzymes at mRNA and activity
levels in pancreas homogenates, and the effects of high and low feed intakes
during the 1st wk postweaning (trial 2) on these measures. The trypsin mRNA
level was transiently decreased 43% 3 d postweaning (P < 0.05). Thereafter,
trypsin and lipase mRNAs linearly increased (P < 0.05). During the 1st wk
postweaning, trypsin- and lipase-specific activities were reduced 44 and 79% (P
< 0.05), respectively, whereas 14 d after weaning, trypsin was at the preweaning
value and lipase was at a low level. Amylase-specific activity did not change
with weaning. Plasma cholecystokinin (CCK) and gastrin concentrations decreased
1 d postweaning and increased afterward up to 3 and 5 d postweaning,
respectively. By 3 d after weaning, the mRNA level of trypsin was twofold higher
(P < 0.05) in piglets that consumed more feed than in those that consumed less,
whereas 7 d after weaning, the groups did not differ. By 7 d after weaning, the
specific activity of amylase was higher, and lipase-specific activity was lower,
in piglets that consumed more feed than in those that consumed less. Plasma CCK
and gastrin concentrations measured 7 d after weaning were correlated with feed
intake (r = +0.56 and r = +0.68, P < 0.05, respectively). In conclusion, by 3 d
postweaning, pancreatic exocrine function was adapting to the new diet.
Afterward, the expression of specific genes coding digestive enzymes and the
levels of pancreatic enzyme activities were restored or stimulated, except for
lipase-specific activity. Therefore, the pancreas can adjust to weaning and dry
food intake as early as wk 2 of life.
Marion et al. J. Nutr 2003; 133:362-368
The effects of intracerebroventricular administration of devezapide, a CCK(1)
receptor antagonist, was investigated on food intake in rats. In the first experiment,
rats (n=5) were deprived of food for 17 h and injected intracerebroventricularly
with either vehicle or devazepide (1, 10, 25 or 100 ng). Five minutes after vehicle
or drug administration, the animals were presented with food and intake measured
for 60 min. Devazepide produced a dose-related increase in food intake. Doses
of 1, 10 and 25 ng significantly increased consumption (at least P<0.01
in each case). A second experiment was subsequently undertaken to investigate
whether systemic administration of the intracerebroventricular doses used in
the first experiment would affect food intake. Rats (n=8) that have been deprived
of food for 17 h were injected intraperitoneally with either vehicle or devazepide
(3, 30, 75 or 300 ng/kg). Five minutes after vehicle or drug administration,
the animals were presented with food and intake was measured for 60 min. Devazepide
(3-300 ng/kg, i.p.) had no significant effects on food consumption. The results
show that central administration of low doses of devazepide increase food intake
in rats, while similar doses, given systemically, do not affect consumption.
These findings suggest the possibility that endogenous cholecystokinin (CCK),
acting at central CCK(1) receptors, may play a physiological role in the control
of feeding behaviour in the rat.
Ebenezer IS. Eur J Pharmacol 2002 Apr 19;441(1-2):79-82
In the present study, we tested the hypothesis that a single daily injection of the gut peptide CCK, together with continuous leptin infusion, would produce
significantly greater loss of body weight than leptin alone. We found that
a single daily intraperitoneal injection of CCK-8 (0.5 microg/kg) significantly
enhanced the weight-reducing effects of 0.5 microg/day leptin infused continuously
into the lateral ventricle of male Sprague-Dawley rats by osmotic minipump.
However, CCK and leptin together did not enhance reduction of daily chow intake.
Furthermore, there was no synergistic reduction of 30-min sucrose intake, although
a significant main effect of both leptin and CCK was observed on sucrose intake.
These results 1) confirm our previous reports of synergy between leptin and
CCK on body weight, 2) demonstrate that enhancement of leptin-induced weight
loss does not require bolus administration of leptin, and 3) suggest that enhanced
body weight loss following leptin and CCK does not require synergistic reduction
of food intake by leptin and CCK.
Matson et al. Am J Physiol Regul Integr Comp Physiol 2002 May;282(5):R1368-73
Type A CCK receptor (CCKAR) antagonists differing in blood-brain barrier permeability [devazepide penetrates; the dicyclohexylammonium salf of Na-3-quinolinoyl-D-Glu-N,N-dipentylaminde (A-70104) does not] were used to test the hypothesis that duodenal nutrient-induced inhibition of gastric emptying is mediated by CCKARs located peripheral to the blood-brain barrier. Rats received A-70104 (700 or 3,000 nmol . kg-1. h-1) or devazepide (2.5 umol/kg iv) and either a 15-min intravenous infusion of CCK-8 (3nmol.kg-1.h-1) or duodenal infusion of casein, peptone, Intralipid, or maltose. Gastric emptying of saline was measured during the last 5 min of each infusion. A-70104 and devazepide abolished the gastric emptying response to a madximal inhibitory dose of CCK-8. Each of the macronutrients inhibited gastric emptying. A-70104 and devazepide attenuated inhibitory responses to each macronutrient. Intravenous injection of a CCK antibody to immunoneutralize circulating CCK had no effect on peptone or Intralipid-induced responses. Thus endogenous CCK appears to act in part by a paracrine or neurocrine mechanism at CCKARs peripheral to the blood-brain barrier to inhibit gastric emptying.
Reidelberger et al. Am J Physiol Regul Integr Comp Physiol 2003; 284: R66-R75
The presence of nutrients in the small intestine slows gastric
emptying and suppresses appetite and food intake; these effects are partly mediated
by the release of gut hormones, including CCK. We investigated the hypothesis
that the modulation of antropyloroduodenal motility, suppression of appetite,
and stimulation of CCK and glucagon-like peptide-1 secretion by intraduodenal
fat are dependent on triglyceride hydrolysis by lipase. Sixteen healthy, young,
lean men were studied twice in double-blind, randomized, crossover fashion. Ratings
for appetite-related sensations, antropyloroduodenal motility, and plasma CCK
and glucagon-like peptide-1 concentrations were measured during a 120-min duodenal
infusion of a triglyceride emulsion (2.8 kcal/min) on one day with, on the other
day without, 120 mg tetrahydrolipstatin, a potent lipase inhibitor. Immediately
after the duodenal fat infusion, food intake at a buffet lunch was quantified.
Lipase inhibition with tetrahydrolipstatin was associated with reductions itonic
and phasic pyloric pressures, increased numbers of isolated antral and duodenal
pressure waves, and stimulation of antropyloroduodenal pressure-wave sequences
(all P < 0.05). Scores for prospective consumption and food intake at lunch were greater, and nausea scores were slightly less, and the rises in plasma CCK and glucagon-like peptide-1 were abolished (all P < 0.05).
In conclusion, lipase inhibition attenuates the effects of duodenal fat on antropyloroduodenal
motility, appetite, and CCK and glucagon-like peptide-1 secretion.
Feinle C et al. Am J Physiol Gastrointest Liver Physiol 284: G798-G807, 2003
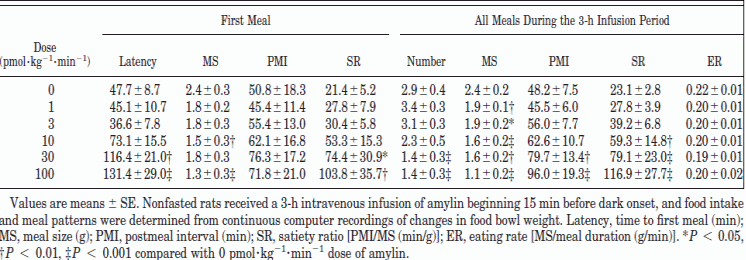
Reidelberger et al. Am J Physiol Regul Integr Comp Physiol 280: R605-R611, 2001
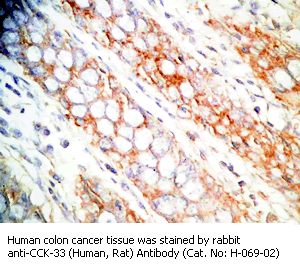 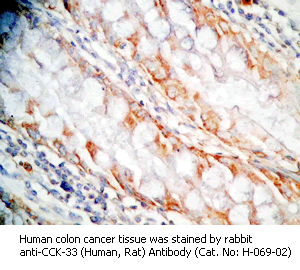
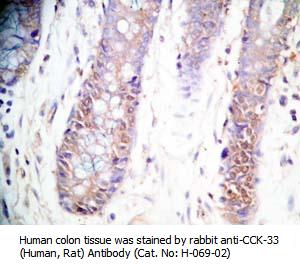 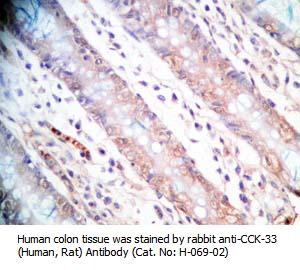
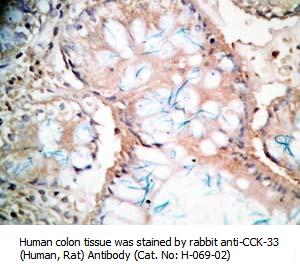
Tissue Sample |
Human colon and colon cancer tissues |
Fixative |
10% formalin |
Embedding |
Paraffin |
Negative Control |
No primary antibody |
Pretreatment |
N/A |
Blocking |
3% H2O2, 2% Normal Goat Serum |
Primary Antibody |
rabbit anti-CCK-33 (Human, Rat) antibody (Cat. No.: H-069-02) |
Optimal Dilution |
1: 500 |
Secondary Antibody |
Goat Anti-Rabbit IgG, Biotinylated (1:400), 30 min |
Amplification |
Streptavidin-HRP (Vector), 1:400, 30 min |
Detection System |
HRP |
Substrate |
DAB (Sigma), 3 min |
Counterstained |
Hematoxylin, 30 sec |
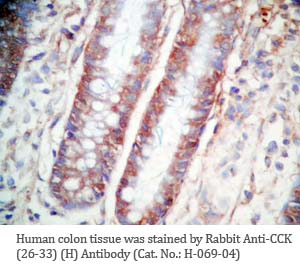 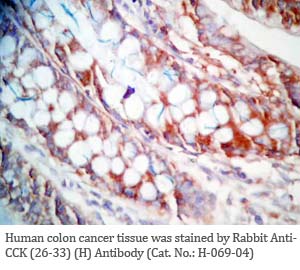
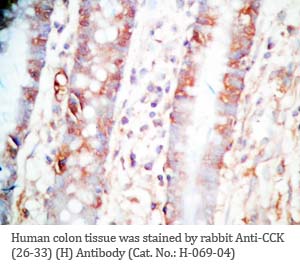 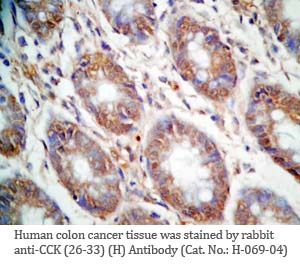
Tissue Sample |
Human colon and colon cancer tissues |
Fixative |
10% formalin |
Embedding |
Paraffin |
Negative Control |
No primary antibody |
Pretreatment |
N/A |
Blocking |
3% H2O2, 2% Normal Goat Serum |
Primary Antibody |
rabbit anti-CCK (26-33) (H,R,M) antibody (Cat. No.: H-069-04) |
Optimal Dilution |
1: 500 |
Secondary Antibody |
Goat Anti-Rabbit IgG, Biotinylated (1:400), 30 min |
Amplification |
Streptavidin-HRP (Vector), 1:400, 30 min |
Detection System |
HRP |
Substrate |
DAB (Sigma), 3 min |
Counterstained |
Hematoxylin, 30 sec |
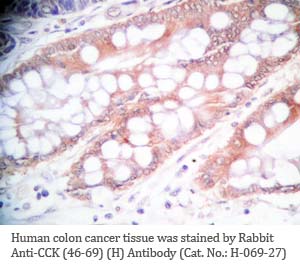
Tissue Sample |
Human colon cancer tissues |
Fixative |
10% formalin |
Embedding |
Paraffin |
Negative Control |
No primary antibody |
Pretreatment |
N/A |
Blocking |
3% H2O2, 2% Normal Goat Serum |
Primary Antibody |
rabbit anti-CCK (46-69) (Human) antibody (Cat. No.: H-069-27) |
Optimal Dilution |
1: 500 |
Secondary Antibody |
Goat Anti-Rabbit IgG, Biotinylated (1:400), 30 min |
Amplification |
Streptavidin-HRP (Vector), 1:400, 30 min |
Detection System |
HRP |
Substrate |
DAB (Sigma), 3 min |
Counterstained |
Hematoxylin, 30 sec |
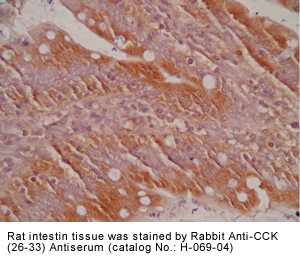
Tissue Sample |
Rat intestine tissues |
Fixative |
10% formalin |
Embedding |
Paraffin |
Negative Control |
No primary antibody |
Pretreatment |
N/A |
Blocking |
3% H2O2, 2% Normal Goat Serum |
Primary Antibody |
rabbit anti-CCK (26-33) (H,R,M) antibody (Cat. No.: H-069-04) |
Optimal Dilution |
1: 500 |
Secondary Antibody |
Goat Anti-Rabbit IgG, Biotinylated (1:400), 30 min |
Amplification |
Streptavidin-HRP (Vector), 1:400, 30 min |
Detection System |
HRP |
Substrate |
DAB (Sigma), 3 min |
Counterstained |
Hematoxylin, 30 sec
|
|
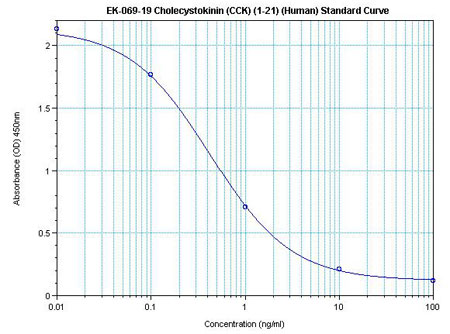
Linear Range: 0.11-2.33 ng/ml
|
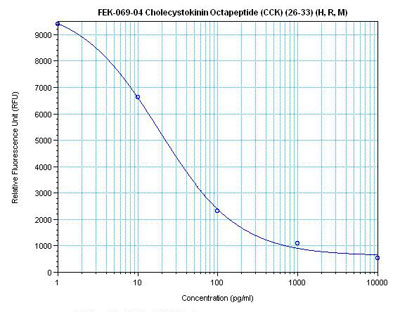
Linear Range 15-333 pg/ml
7 times more sensitive than normal EIA kits
|

Linear Range: 2.36 - 90 pg/ml
Over 47 times more sensitive than normal EIA kits | 
Linear Range: 0.11-2.33 ng/ml
|
CCK EIA Kit References
CCK RIA Kit References
|
|
|
%CCK%;%069-10%;%069-08%
|
|
|




-(Human)-Standard-Curve.jpg)
-Standard-Curve.jpg)