|
Prolactin-releasing Peptide (PrRP)
Prolactin
Releasing Peptides (PrRP) Related Products



| |
Alternative role for prolactin-releasing peptide in the regulation
of food intake.
Catherine B. Lawrence, et al.
Nature Neuroscience 3, 645 - 646 (2000)
|
|
PrRP affects food intake and core body temperature in free-fed
animals.
(a) Cumulative food intake was significantly reduced after
4 nmol PrRP (i.c.v.; * p < 0.05 versus vehicle). (b) Temperature
is expressed as change in core body temperature from baseline
(time zero) and was measured by remote telemetry. 2 nmol or
4 nmol PrRP caused a hypothermic response that reached a nadir
at 30 min and then returned to above control levels at 2 h
after injection. Rat PrRP-31 (Bachem, Saffron Walden, UK)
was injected through indwelling catheters into the lateral
ventricle of conscious, free-moving, male Sprague-Dawley rats
(n = 6-7 per group). Food and water (see text) intake data
were analyzed using repeated measures ANOVA with Scheffe's
post-hoc comparisons. Otherwise, analyses used a one-way ANOVA
with Tukey post-hoc comparisons.
Catherine B. Lawrence, et al.
Nature Neuroscience 3, 645 - 646 (2000)
|
|
|
PrRP reduces fast-induced re-feeding.
Animals were allowed to re-feed after a 24-h fast. (a) 4 nmol
PrRP significantly reduced the initial re-feeding response,
though food intake was normalized in PrRP-treated animals
during the 24 h after injection. (b)Although core body temperature
is strongly affected by fast-induced re-feeding, the change
in body temperature was consistently above that of controls
for a period of several hours. Statistics involved repeated
- measures ANOVA and t-tests. * p < 0.05, ** p < 0.01
and *** p < 0.001 versus vehicle-treated group.
Catherine B. Lawrence, et al.
Nature Neuroscience 3, 645 - 646 (2000)
|
|
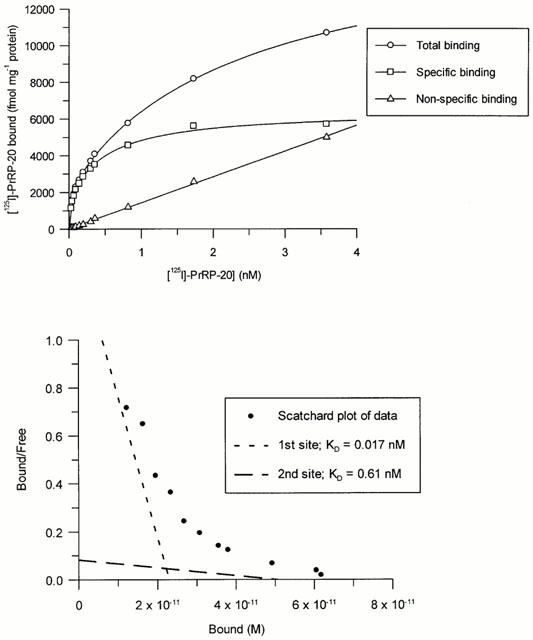
(a) Total, non-specific and specific binding of [125I]-PrRP-20
to membranes from HEK293 cells expressing GPR10 receptors
with increasing radioligand concentration. Data represents
a single experiment (each point determined in quadruplicate),
which was replicated six times with similar results. (b) Scatchard
transformation of the data from (a).
Langmead C.L. et al. British
Journal of Pharmacology (2000) 131, 683-688
|
|
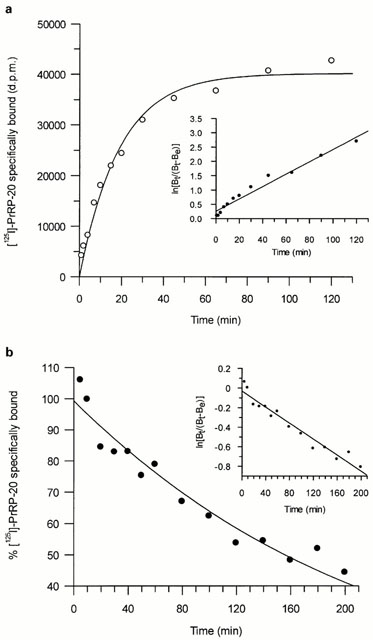
(a) Time course for association of [125I]-PrRP-20 binding
to HEK293-GPR10 receptor expressing membranes. Data represents
a single experiment, which was replicated five times with
similar results. The inset shows the data transformed as a
semi-log plot (correlation coefficient r=0.98), where Bt is
the specific binding at time t and Be is the specific binding
measured at equilibrium. (b) Time course for the dissociation
of [125I]-PrRP-20 binding from HEK293-GPR10 receptor expressing
membranes. Data represents a single experiment, which was
replicated three times with similar results. The inset shows
the data transformed as semi-log plot, where Bt is the specific
binding at time t and Be is the specific binding measured
at equilibrium.
Langmead C.L. et al. British
Journal of Pharmacology (2000) 131, 683-688
|
|
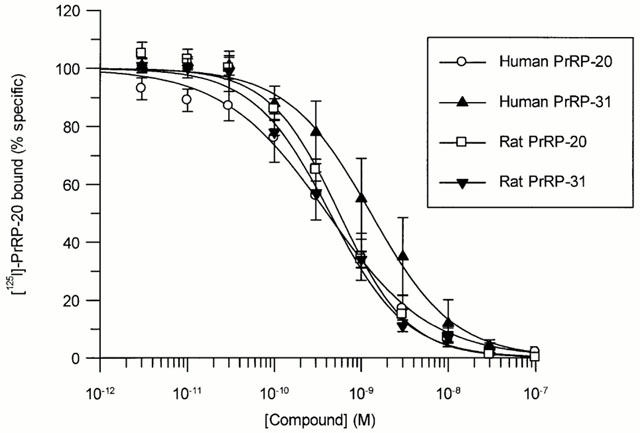
Competition for [125I]-PrRP-20 binding to HEK293-GPR10 expressing
membranes by rat and human PrRP-20 and PrRP-31. [125I]-PrRP-20
(0.2 nM) was incubated in the presence of increasing concentrations
of the compounds. Data are the mean of at least three independent
experiments; vertical lines show s.e.mean.
Langmead C.L. et al. British
Journal of Pharmacology (2000) 131, 683-688
|
|
|
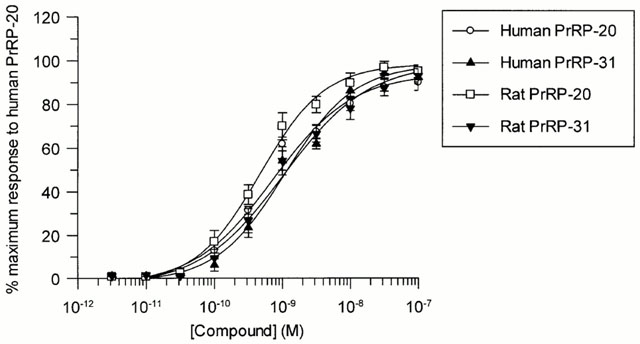
Concentration dependent stimulation of Ca2+ mobilization by
human PrRP-20, human PrRP-31, rat PrRP-20 and rat PrRP-31
in HEK293 cells expressing GPR10. Data shown are the mean
of six experiments; vertical lines show s.e.mean.
Langmead C.L. et al. British
Journal of Pharmacology (2000) 131, 683-688
|
|
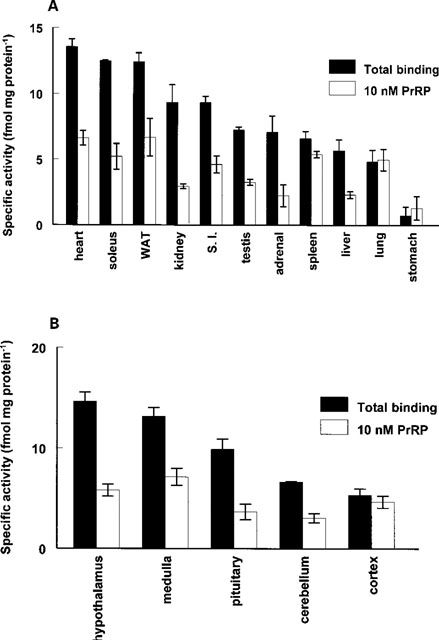
The distribution of 125I-PrRP binding in rat peripheral tissues
(A) and the central nervous system (B) in the presence or
absence of 10 nM hPrRP. Rat tissue membranes (100 g membrane
protein) were prepared and binding assays using 125I-hPrRP
(750 Bq, 30 pM) performed as described in the Methods. Binding
is presented as fmols of specific 125I-hPrRP binding per mg
protein. All binding assays were performed in triplicate in
the presence and absence of 1 M hPrRP to calculate specific
binding (labelled total binding, filled bars). Specific binding
was also measured in the presence of 10 nM hPrRP (open bars).
Each time point is the means.e.mean of at least three separate
membrane preparations. WAT is white adipose tissue; S.I. is
small intestine; medulla is medulla oblongata; cortex is cerebral
cortex.
Fumitoshi Satoh, et al. British
Journal of Pharmacology (2000) 129, 1787-1793
|
|
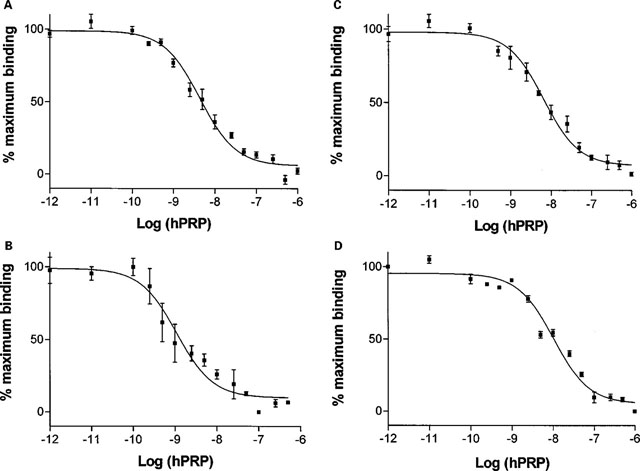
Competition for 125I-PrRP binding by hPrRP in rat hypothalamic
(A), pituitary (B), heart (C) and soleus muscle (D) membranes.
Rat tissue membranes (100 g membrane protein) were prepared
and binding assays using 125I-hPrRP (750 Bq, 30 pM) performed
as described in the Methods. Non-specific binding was measured
in the presence of 1 M hPrRP. Binding is shown as a percentage
of the maximal specific binding in the absence of unlabelled
PrRP. All binding assays were performed in triplicate and
the curves shown are meanss.e.mean of five (hypothalamus),
four (heart and soleus) and three (pituitary) separate experiments.
Fumitoshi Satoh, et al. British
Journal of Pharmacology (2000) 129, 1787-1793
|
|
Amino acid sequences of short and long forms of prolactin
releasing peptide (PrRP) in rat and human.Langmead C.L.
et al.
British Journal of Pharmacology
(2000) 131, 683-688
|
|
Membrane preparation |
Human embryonic kidney cells (HEK293) stably transfected
with GPR10 were harvested with PBS, pelleted and stored at
-80°C until further use. Membranes were prepared using a modification
of the method of Miyamoto et al. (1994); all procedures were
carried out at 4°C. In brief, cells were washed in 30 vols
(w v-1) of PBS with 0.2 mM EDTA. The suspension was homogenized
using an Ultra-Turrax homogenizer and the subsequent homogenates
centrifuged at 39,000g for 15 min. The resultant pellets were
resuspended in 30 volumes of buffer containing 10 mM Na2CO3,
1 mM EDTA, 0.5 mM phenylmethylsulphonyl fluoride (PMSF), 1
g ml-1 pepstatin and 1CompleteTM serine and cysteine protease
inhibitor tablet 250 ml-1 (pH 7.4). The suspension was then
homogenized and centrifuged at 1000g for 10 min, the supernatant
decanted and centrifuged at 48,000g for 20 min. The resultant
pellets were resuspended in buffer containing 20 mM Tris-HCl,
0.25 M sucrose, 2 mM EDTA, 0.5 mM PMSF, 1 g ml-1 pepstatin
and 1CompleteTM serine and cysteine protease inhibitor tablet
250 ml-1 (pH 7.4) to a volume of approximately 48106 cells
ml-1 and stored at -80°C until used.
Langmead C.L. et al. British
Journal of Pharmacology (2000) 131, 683-688
|
| [125I]-PrRP-20 binding assays |
HEK293-GPR10 receptor expressing cell membranes were incubated
with [125I]-PrRP-20 in buffer containing 20 mM Tris-HCl, 5
mM Mg-Acetate, 2 mM EGTA, 0.5 mM PMSF, 1 g ml-1 pepstatin
and 1CompleteTM serine and cysteine protease inhibitor tablet
250 ml-1 and 0.1% (w v-1) BSA (pH 7.4) at 25°C for 90 min.
The total assay volume was 0.5 ml. The reaction was terminated
by rapid filtration through Whatman GF/B glass fibre filters,
followed by rapid washing of the filters with 51 ml aliquots
of ice cold buffer containing 50 mM Tris-HCl, 10 mM MgCl2
(pH 7.4). Bound radioactivity was determined by gamma counting.
Non-specific binding was defined as that remaining in the
presence of 0.1 M rat PrRP-31. Saturation studies were carried
out by incubating membranes (4 g protein well-1) with a range
of concentrations of [125I]-PrRP-20 (0.01-5 nM). Specific
binding data was analysed using the program Radlig (Biosoft)
to provide estimates of KD and Bmax values. Protein content
was assayed using the Bradford method (Bradford, 1976) using
bovine serum albumin as a standard. Association kinetic studies
were performed by measuring specific binding of [125I]-PrRP-20
(0.2 nM) at 0.5-90 min after addition of membranes (2 g protein
well-1). For dissociation studies, membranes were pre-incubated
with [125I]-PrRP-20 (0.2 nM) for 90 min. Specific binding
was then measured at 5-200 min after the addition of 0.1 M
PrRP-31. Kinetic data was analysed by GraFit (Erithacus Software)
to provide estimates of Kon and Koff values. Competition studies
were performed by incubating cell membranes (2 g protein well-1)
with [125I]-PrRP-20 (0.2 nM) and a range of concentrations
of the test compound. Competition curves were analysed by
non-linear least-squares fitting to a four parameter logistic
equation by Microsoft Excel in order to determine IC50 values
(Bowen & Jerman, 1995). Ki values were then derived from
the IC50 values using a nominal KD value of 0.1 nM (which
takes into account binding to both high and low affinity sites
obtained from saturation studies) (Cheng & Prussoff, 1973).
Results are given as means (s.e.mean) of at least three independent
experiments.
Langmead C.L. et al. British
Journal of Pharmacology (2000) 131, 683-688
|
| Calcium mobilization assays |
Intracellular calcium was monitored using the fluorescent
dye Fluo 4AM in a Fluorometric Imaging Plate Reader (FLIPR,
Molecular Devices, U.K.). HEK293-GPR10 cells were cultured in
poly-D-lysine coated 96-well microtitre plates 24 h before use
at as a seeding density of 52,000 cells well-1. Prior to assay
on FLIPR, cells were incubated with Fluo 4AM (1 M) for 60 min
at 37°C in Hank's buffered saline solution containing 0.1% BSA
and 2.5 mM probenicid. Extracellular dye was then removed by
washing three times with 150 l Hank's buffered saline solution
containing 2.5 mM probenicid without BSA. Compounds were tested
for agonist activity in FLIPR by adding 40 l of test solution
to a plate volume of 120 l at 37°C. Peak stimulation (minus
basal) was plotted versus concentration of test compound and
iteratively curve fitted using a four parameter logistic equation
(Grafit, Erithacus Software) to assess agonist potency and maximal
response.
Langmead C.L. et al. British Journal
of Pharmacology (2000) 131, 683-688 |
| |
| |
| |
1. Lawrence CB, Celsi F, Brennand J,
Luckman SM. Alternative role for prolactin-releasing peptide in the
regulation of food intake. Nat Neurosci 2000 Jul;3(7):645-646 School
of Biological Sciences, University of Manchester, Oxford Road, Manchester
M13 9PT UK.
1. Prolactin-releasing peptide (PrRP) is a peptide ligand for the human
orphan G-protein-coupled receptor hGR3/GPR10 and causes the secretion
of prolactin from anterior pituitary cells. However, the lack of immunoreactive
staining for PrRP in the external layer of the median eminence seems
to rule out this peptide as a classical hypophysiotropic hormone and,
furthermore, PrRP is less effective than another inducer of prolactin
secretion, thyrotropin-releasing hormone, both in vitro and in vivo.
Here we show a reduction in the expression of PrRP mRNA during lactation
and fasting and an acute effect of PrRP on food intake and body weight,
supporting the hypothesis of an alternative role for the peptide.
2. Samson WK, Resch ZT, Murphy TC. A novel action of the newly described prolactin-releasing
peptides: cardiovascular regulation. Brain Res 2000 Mar 6;858(1):19-25.
3. Satoh F, Smith DM, Gardiner JV, Mahmoodi M, Murphy KG, Ghatei MA,
Bloom SR
Characterization and distribution of prolactin releasing
peptide (PrRP) binding sites in the rat--evidence for a novel binding
site subtype in cardiac and skeletal muscle. Br J Pharmacol 2000
Apr;129(8):1787-93
Endocrine Unit of the Department of Metabolic Medicine, Imperial College
School of Medicine, Hammersmith Hospital, London, W12 ONN.
Prolactin releasing peptide (PrRP) was recently purified from bovine hypothalamus
and binds to the orphan receptor, UHR-1. We examined the distribution
and kinetics of (125)I-PrRP binding in rat tissues together with molecular
characterization by chemical cross-linking and Northern blotting. In this
study (125)I-PrRP binding showed specificity and rapid association and
dissociation. Specific binding was found in membranes from rat tissues
including brain (hypothalamus, medulla oblongata and cerebellum), pituitary,
heart, soleus muscle, adipose tissue, kidney, adrenal gland, testis and
small intestine. In hypothalamus, pituitary, heart and soleus competition
analysis indicated only one class of binding site in each tissue. Binding
affinity for PrRP (IC(50)) and binding site density (B(max)) respectively
were 5.2+/-0.9 nM and 674+/-97 fmol mg protein(-1) in hypothalamus (n
= 5), 1.4+/-0.6 nM and 541+/-126 fmol mg protein(-1) in pituitary (n =
3), 6.6+/-0.7 nM and 628+/-74 fmol mg protein(-1) in heart (n = 4) and
9.8+/-0.9 nM and 677+/-121 Soun mg protein(-1) in soleus muscle (n = 4).
Analysis of (125)I-PrRP-binding site complexes by chemical cross-linking
showed a binding site M(r) of 69,000 in hypothalamus and 41,000 in heart
and soleus. Northern analysis of polyA(+) RNA from hypothalamus showed
a 4.2 kb band as expected for UHR-1, but heart and soleus showed a 4.8
kb band. Taken together these results indicate that there may be different
subtypes of PrRP binding sites in rat tissues which may differ from UHR-1.
4. Matsumoto H, Murakami Y, Horikoshi Y, Noguchi J, Habata
Y, Kitada C, Hinuma S, Onda H, Fujino M. Distribution and characterization
of immunoreactive prolactin-releasing peptide (PrRP) in rat tissue and
plasma. Biochem Biophys Res Commun 1999 Apr 13;257(2):264-8
We established a sensitive and specific two-site enzyme immunoassay
(EIA) for prolactin-releasing peptide (PrRP) using two region-specific
monoclonal antibodies. We investigated the tissue distribution and the
plasma concentration of immunoreactive (ir-) PrRP in rats using this
assay. Ir-PrRP was widely distributed in the central nervous system
and pituitary gland. The highest concentration of ir-PrRP was found
in the hypothalamus. In peripheral tissues, appreciable levels of ir-PrRP
were found only in the adrenal gland. The mean plasma concentration
of ir-PrRP was 0.13 +/- 0.01 fmol/ml (mean +/- SEM). In reverse-phase
and gel-filtration high performance liquid chromatography, hypothalamic
ir-PrRP eluted at a position identical to that of PrRP31 and PrRP20.
On the other hand, ir-PrRP from the adrenal gland and plasma eluted
only at the position of synthetic PrRP31, indicating that molecular
forms of ir-PrRP in vivo differed among tissues. Copyright 1999 Academic
Press.
The prolactin-releasing peptide receptor (GPR10) regulates
body weight homeostasis in mice.
To identify new drug targets for the treatment of obesity, we employed
a degenerate reverse transcriptasepolymerase chain reaction technique
to isolate novel members of the G-protein coupled receptor superfamily
from mouse hypothalamus. One of our clones was found to encode a protein
with 90% amino acid identity to human GPR10, which was previously identified
as the receptor for prolactin-releasing peptide (PrRP) and has been
implicated in lactation, the regulation of food intake and other physiological
functions. To investigate the role of GPR10 in food intake and body
weight homeostasis, we generated mice carrying a targeted deletion of
the GPR10 gene. First, using these knockout animals, we confirmed that
GPR10 is the principle receptor for PrRP in the mouse hypothalamus because
deletion of GPR10 completely abolished PrRP binding to isolated hypothalamic
cell membranes. Second, we investigated the effect of normal and high-fat
diets on energy intake, body weight, and glucose homeostasis in wild-type
and GPR10 knockout mice. After fasting and refeeding, food intake in
knockout animals was unchanged relative to control littermates. However,
beginning at 16 wk of age on a normal diet, knockout mice became hyperphagic,
obese, and showed significant increases in body fat and the levels of
leptin and insulin, as well as decreased glucose tolerance. This metabolic
profile was similar to the effect of a high-fat diet on wild-type animals.
Our findings provide direct evidence that GPR10 is the receptor for
PrRP and that it is involved in the regulation of energy balance in
mice. GPR10 knockout mice will also prove useful for investigating other
proposed activities for PrRP.
Gu W, et al. J Mol Neurosci. 2004;22(1-2):93-103
PrRP
Related Products
|